
Relation structure fonction
Les globines constituent une famille de protéines largement répandues. Elles sont toutes impliquées dans le transport ou le stockage du dioxygène. L'hémoglobine HBA (hémoglobine adulte normale), dissoute dans le cytosol aqueux des érythrocytes en une solution très concentrée, assure le transport du dioxygène des poumons vers les tissus. Un globule rouge humain de 7 mm, contient 280 millions de molécules d’hémoglobine.
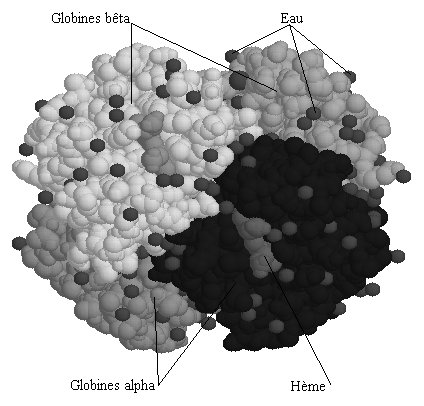
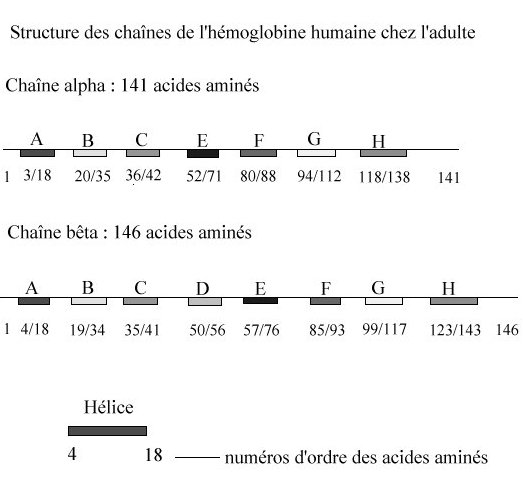
Chaque molécule d’hémoglobine est un tétramère formé par l’association de quatre chaînes polypeptidiques identiques deux à deux : deux chaînes alpha (ou globines alpha) composées chacune de 141 acides aminés et deux chaînes bêta (ou globines bêta) de 146 acides aminés. Chacune des chaînes adopte une conformation spatiale lui donnant une forme globuleuse et ménageant une « poche » superficielle dans laquelle se trouve logé le hème. La cohérence du tétramère (structure quaternaire de l’hémoglobine a2-b2) résulte de liaisons dues aux chaînes latérales hydrophobes des acides aminés situés à la périphérie de chaque globine.
Une chaîne polypeptidique alpha ou bêta présente sept ou huit secteurs en forme d’hélice droite reliés par des segments comportant parfois des coudes.
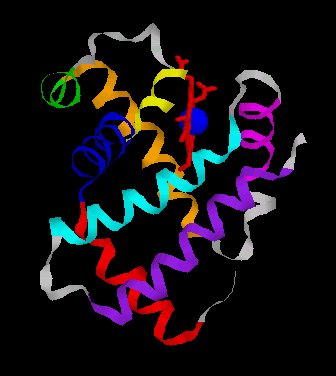 | Images d'une globine beta : A gauche, une globine en ruban colorée par hélice (hème en rouge, fer en bleu); A droite, un schéma de cette globine
|  |
Bien que les chaînes alpha et bêta aient des séquences différentes, elles présentent des structures tertiaires assez similaires.
 | Globine alpha 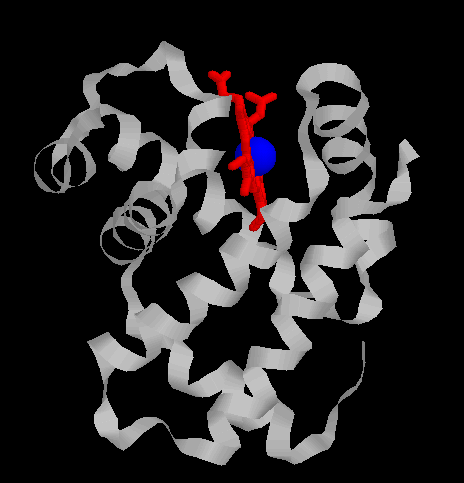 Globine beta 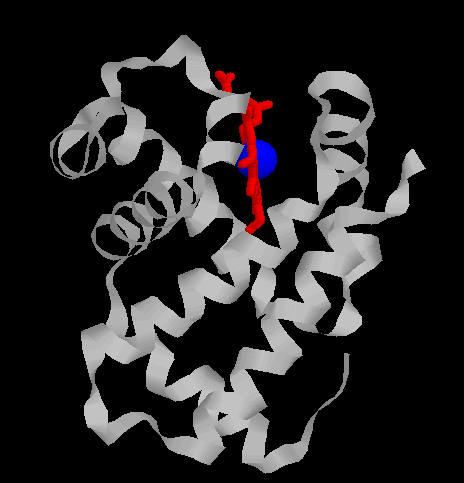 |
Quatre sous-unités s'associent dans une symétrie tétraédrale pour former la molécule d'hémoglobine (fichier 3D de 50 Ko - molécule partielle restreinte à ses seuls carbones alpha), structure globulaire de 65 x 50 x 55 Å. En fait, le tétramère résulte de l'association de deux dimères fonctionnels : a1 b1 et a2 b2. Dans la structure tétramérique, les dimères sont disposés de façon à ce que la sous-unité a1 soit au contact de la sous-unité b2 et a2 de b1.
La disposition des chaînes est telle que des rapports très intriqués existent entre chaînes latérales de résidus appartenant aux sous-unités non homologues. A l'inverse, il n'existe qu'un faible nombre de contacts entre sous-unités identiques.
Trois zones de contact sont donc à distinguer :
- Contacts entre sous-unités d'un même dimère (a1 b1 ou a2 b2)
- Contacts entre chaînes non homologues de deux dimères différents (a1 b2 ou a2 b1)
- Contacts entre chaînes homologues : Le plus important d'entre eux est établi entre les chaînes b, au niveau de la cavité centrale, par l'intermédiaire d'une molécule de 2,3 diphosphoglycérate (2,3-DPG), et stabilise la configuration désoxygénée.
C'est au niveau de cette zone que s'effectuent les mouvements de glissement et de rotation qui accompagnent la modification de conformation de l'hémoglobine lors de l'oxygénation. Les résidus en contact diffèrent pour chacune des configurations.
Dans la désoxyhémoglobine, des interactions électrostatiques faisant intervenir l'ion Cl- s'établissent également entre l'extrémité C-terminale d'une chaîne a et l'extrémité N-terminale de l'autre.
Chez tous les vertébrés, l'hémoglobine est un tétramère dont les sous-unités sont identiques deux à deux et se distinguent en type alpha et béta. La structure spatiale de ces sous-unités a été remarquablement conservée (2 fichiers 3D de 14 Ko chacun - molécules restreintes aux seuls carbones alpha) depuis l'apparition de la molécule ancestrale qui remonte sans doute à quelques 600 à 700 millions d'années. Une duplication ultérieure du gène hémoglobinique a conduit aux deux types de chaînes a et b et à l'apparition de l'hétéro-tétramère. A partir de ce stade, des duplications successives, l'accumulation de mutations et la pression sélective ont conduit à des molécules parfaitement adaptées aux diverses conditions d'environnement. Les études par diffraction de rayons X ont permis dès 1960 de connaître la structure spatiale de la molécule d'hémoglobine avec une résolution de 5,5 Å. L'interprétation de ces résultats a grandement bénéficié de ceux obtenus à plus forte résolution pour la myoglobine. Aujourd'hui, la structure spatiale de l'hémoglobine est connue avec un degré de résolution de 1,7 Å, ce qui donne une excellente définition de la position de chaque atome. La comparaison des structures primaires des hémoglobines monomériques (ou des sous-unités), depuis les espèces animales les plus primitives jusqu'à l'homme, est assez décevante : les homologies entre espèces éloignées sont faibles, et à priori rien ne laisserait même penser à une parenté structurale. Au contraire, la comparaison des structures spatiales montre une géométrie remarquablement conservée.
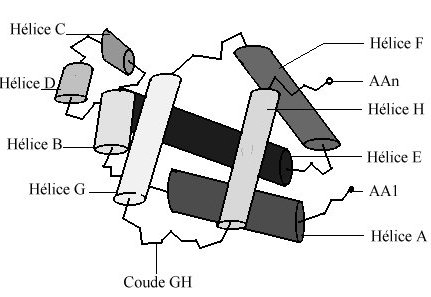
Toutes les myoglobines et sous-unités d'hémoglobines, qu'elles proviennent des leghémoglobines des rhizomes (du règne végétal) ou des hémoglobines (de la série animale) partagent une même structure spatiale. Chaque globine est caractérisée par la présence de huit segments hélicoïdaux , repliés de façon à ménager une cavité hydrophobe dans laquelle une molécule d'hème est enfouie (attention fichier de 78 Ko - molécule beta complète).
Les sous-unités d'hémoglobine peuvent être assimilées à des micelles : leur surface externe, en contact avec le milieux aqueux ambiant, est tapissée par les chaînes latérales de résidus hydrophiles ; les régions internes sont, quant à elles, essentiellement occupées par des résidus hydrophobes. Ces derniers, en échangeant un très grand nombre de liaisons de faible énergie, stabilisent l'édifice moléculaire. Pour faciliter la comparaison entre sous-unités, l'usage est d'utiliser la nomenclature stéréochimique. Les hélices sont ainsi désignées par une lettre de A à H, en partant de l'extrémité N terminale ; les segments inter-hélicoïdaux par les noms des hélices qui leur sont adjacentes ; les résidus par leur position dans ces éléments structuraux. Dans cette nomenclature, l'histidine proximale est toujours en position F8 alors qu'en appliquant la nomenclature linéaire, elle occupe la position a87 ou b 92, selon qu'il s'agit de la chaîne a ou b.
La cavité en forme de V, dans laquelle la molécule d'hème est logée, est limitée par les hélices B, C, E, F, G, H et par le segment CD.
| L'Hème |
L’oxygénation de l’hémoglobine provoque une modification de sa conformation tridimensionnelle : la molécule d’oxyhémoglobine est un peu plus compacte que celle de la désoxyhémoglobine ; les hèmes subissent des modifications spatiales lors de la fixation du dioxygène, l’atome de fer se déplace.
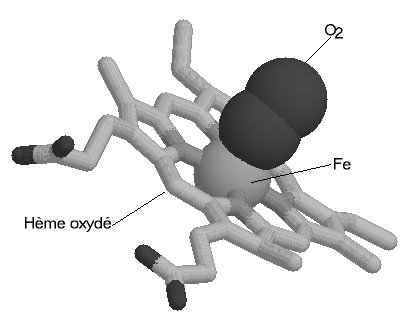
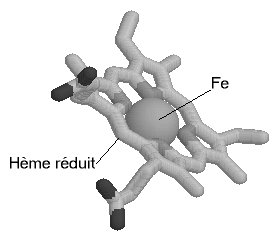
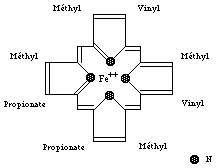
Une molécule d'hème (fichier de 14 Ko - molécule restreinte aux seuls carbones alpha) est liée à chacune des sous-unités. Il s'agit d'une molécule plane, ou légérement bombée, selon le groupe lié à sa sixième valence de coordinence. Sa structure est connue depuis le début du siècle : elle est constituée par une protoporphyrine ayant à son centre un atome de fer. La protoporphyrine est formée de quatre cycles pyrroliques unis par l'intermédiaire de ponts méthénylés (-CH=) et substitués par des groupes méthyl, propionate et vinyl dont la disposition est caractéristique du type IX . Que la molécule d'hémoglobine soit oxygénée (oxyhémoglobine) ou désoxygénée (désoxyhémoglobine), le fer reste sous forme réduite (Fe++) (2 fichiers de 14 Ko chacun - molécules restreintes aux seuls carbones alpha).
Dans l'oxyhémoglobine, l'atome de fer présente six liaisons de coordinence : quatre interviennent dans la structure de l'hème, la cinquième amarre l'hème à la globine au niveau de l'histidine F8 (histidine proximale) et la sixième fixe une petite molécule tel l'oxygène, appelé "ligand". Ce ligand est en rapport avec l'histidine E7 (histidine distale). Dans la désoxyhémoglobine, où aucun ligand n'occupe la face distale de l'hème, le fer est pentacoordonné. Par suite d'une distribution différente des électrons dans les couches périphériques, le volume de cet atome augmente. Ces changements de taille sont à la base même des mécanismes de modification de configuration de la structure protéique accompagnant la fixation d'oxygène.
Le rôle de la protéine entourant l'hème peut très schématiquement se résumer à :
- une mise en position optimale de la molécule d'oxygène,
- une protection contre l'oxydation,
- une modulation de la fonction oxyphorique.
L'oxygénation modifie la structure de la molécule. La fixation d'une molécule d'oxygène sur une sous-unité entraine des modifications au niveau de la poche de l'hème, qui se répercutent sur l'extrémité C-terminale de la chaîne b et ainsi sur les contacts entre sous-unités. Un mouvement de glissement et de rotation se produit ainsi au niveau de l'interface a1b2. La conformation T (ou Tense) correspond à la forme désoxygénée et la conformation R (Relax) à la forme oxygénée.
Lors de la transition de la configuration désoxygénée vers la configuration oxygénée, le 2,3-DPG est expulsé de la cavité centrale.