
Polydactylie - Application pédagogique
1 - Présentation de la polydactylie préaxiale
Photos de polydactylie
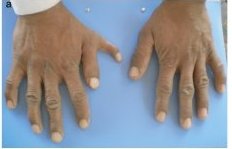
Main d'une personne polydactyle (à gauche) et d'une personne non polydactyle (à droite).
Muhammad Farooq and ali. Preaxial polydactyly/triphalangeal thumb is associated with changed transcription factor-binding affinity in a family with a novel point mutation in the long-range cis-regulatory element ZRS. Eur J Hum Genet. 2010 June; 18(6): 733–736.
Familles avec polydactylie
 |
 |
Les symboles des personnes polydactyles sont en noir.
Lettice LA, Heaney SJH, Purdie LA, Li L, de Beer P, Oostra BA, Goode D, Elgar G, Hill RE and de Graaff E (2003) “A long-range Shh enhancer regulates expression in the developing limb and fin and is associated with preaxial polydactyly” Human Molecular Genetics 12
(14):1725-35.
Ces documents permettent de présenter la polydactylie et de préciser la variante polydactylie préaxiale dont la fréquence est de l’ordre de 1 pour 2000 naissances.
En utilisant le raisonnement vu en classe de première (calcul du risque relatif) on peut établir que le phénotype polydactyle est héréditaire. Il s’agit ensuite de déterminer le modèle génétique capable de rendre compte des caractéristiques de la transmission du phénotype polydactyle dans ces familles. Les données généalogiques s’interprètent bien en admettant que la différence phénotypique est due à un seul gène et que le phénotype polydactyle est dominant. On arrive à l’idée que les individus polydactyles de ces familles sont hétérozygotes pour ce gène.
2 – Le modèle murin de la polydactylie et l’implication du gène sonic hedgehog. (Shh)
Le phénotype polydactyle étant aussi connu chez la souris, l’étude de certains aspects du développement des membres chez l’embryon de souris a permis d’élucider les mécanismes à l’origine de l’anomalie des doigts.
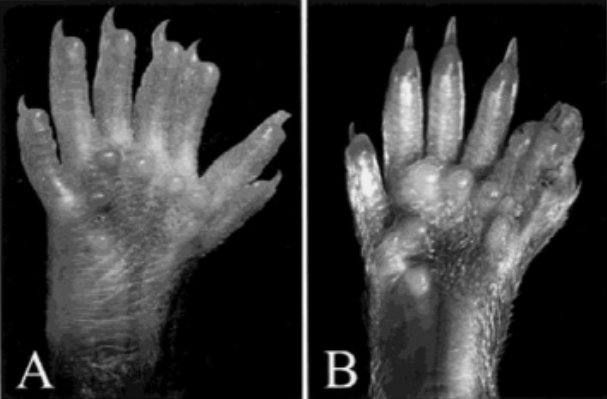
Vue ventrale de membres postérieurs de souris hétérozygote Hx montrant une polydactylie préaxiale.
Le gène Shh et le développement normal du membre
1 - Etapes du développement du membre

Développement embryonnaire d'une patte avant de souris. Les valeurs en bas de chaque image indiquent le nombre de jours après le début de la gestation.
Les photos sur le développement des membres montrent que ceux-ci se forment à partir de petites protubérances, les bourgeons des membres, qui font saillie sur la paroi du corps de l’embryon. Cette formation débute à 9,5 jours de gestation pour les membres avant, 0,5 jour plus tard pour les membres arrière. Les doigts sont esquissés à 12,5j pour le membre avant et à14,5j, un membre en miniature est bien formé.
2 - Axes du développement

Image d'un embryon de souris à 10,5j de gestation. Le zoom montre le bourgeon d'un membre avant avec ses deux axes principaux de développement.
Pour décrire le développement du membre, les embryologistes reconnaissent trois axes (fig.) : l’axe proximo distal (de la base du membre à son extrémité), l’axe antéro-postérieur (la région antérieure correspondant à celle du pouce ou du gros orteil (doigt 1) la région postérieure à celle du doigt 5), l’axe dorso ventral.
3 - Importance de la zone ZPA et du gène Shh
Les études expérimentales ont montré l’importance d’une zone située dans la région postérieure du bourgeon du membre pour l’induction du pattern des doigts. Cette zone est appelée ZPA (zone d’activité polarisante). Elle induit la formation de doigts par l’intermédiaire d’une molécule de signalisation, la protéine sonic hedgehog (Shh) qu’ont produite ses cellules.
4 - Les arguments concernant l'importance de la ZPA et de Shh
A - Expérience de greffe chez le poulet
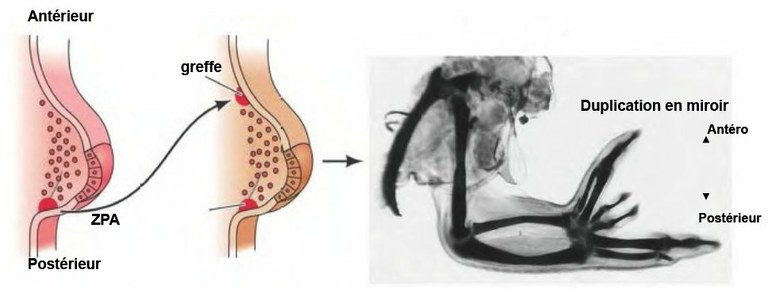
Photo de greffe de la zone ZPA dans la région antérieure du bourgeon du membre chez le poulet. PDF "Faire un membre".
Quand cette ZPA est prélevée sur un jeune bourgeon d’aile de poulet (avant qu’apparaissent les doigts) et greffée sur le bord antérieur d’un autre bourgeon alaire, une aile en miroir se développe : au lieu de la disposition normale des doigts : 4, 3,2 (il n’y a que 3 doigts dans la main du poulet), on obtient le schéma 4, 3, 2, 2, 3,4. Autrement dit le greffon a déterminé la formation de 3 doigts supplémentaires. On a montré que ces 3 doigts ne proviennent pas du greffon mais du bourgeon sur lequel on a réalisé la greffe.
B - Conséquence de la délétion du gène Shh sur le développement de la souris. ChiangNature.
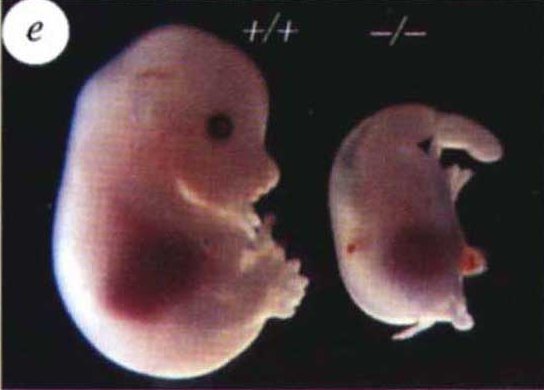
La figure montre l’aspect d’un embryon de souris mutant avec Shh délété par rapport à un embryon normal. La morphologie des membres est anormale (un seul doigt, le doigt 1) mais d’autres régions du corps sont atteintes, notamment le cerveau postérieur et le crâne, montrant que le gène Shh intervient dans différentes régions du corps au cours du développement (les embryons ne sont pas viables).
Etude de l'expression du gène Shh au cours du développement du membre
Les chercheurs ont établi les caractéristiques de l'expression du gène Shh dans le bourgeon du membre (lieu et moment d'expression). Pour cela ils ont mis en évidence l'ARNm spécifique de ce gène par la méthode de l'hybridation in situ. La présence de l'ARNm se traduit par une coloration violette.
Dans tous les clichés qui suivent, la région proximale du membre est à gauche, la région antérieure vers le haut (et donc la région postérieure vers le bas).
Cette expression commence à 9,5j.
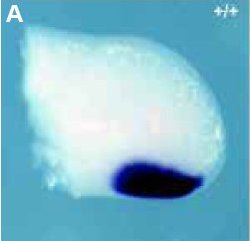
Expression du gène Shh dans le bourgeon du membre au jour 11,5 chez l'embryon de souris normale. La région antérieure est en haut.
mouse mutant Hemimelic extra-toes. ISABELLE BLANC1, ANTOINE BACH and BENOÎT ROBERT. Laboratoire de Génétique Moléculaire de la Morphogenèse, CNRS URA 1947, Institut Pasteur, Paris, France

Expression du gène Shh dans le bourgeon du membre au jour 12 jours chez l'embryon de souris normale.
Unusual pattern of Sonic hedgehog expression in the polydactylous
mouse mutant Hemimelic extra-toes. ISABELLE BLANC1, ANTOINE BACH and BENOÎT ROBERT. Laboratoire de Génétique Moléculaire de la Morphogenèse, CNRS URA 1947, Institut Pasteur, Paris, France
On constate que l'expression de Shh à 11,5j est limitée à la région postérieure du bourgeon dans une zone qui correspond à la ZPA; Cela confirme que cette ZPA peut être impliquée dans la construction des doigts par l'intermédiaire de la protéine Shh.
A 12j on ne distingue plus de coloration violette ce qui indique que le gène a cessé de s'exprimer.
Cette analyse montre qu'au cours de la construction d'un organe, un gène de développement s'exprime pendant un temps donné et dans un territoire précis.
Le gène Shh et la polydactylie
Ces informations sur le développement du membre permettent de suspecter l’implication du gène Shh dans la polydactylie.Les données suivantes permettent de tester cette hypothèse.
A - Comparaison des séquences codantes de Shh de la souris normale et de la souris polydactyle
Ces séquences sont fournies pour une comparaison avec Anagène qui révèle qu'elles sont identiques. Ces résultats ne vont pas dans le sens de l'hypothèse émise.
B - Expression du gène Shh dans les bourgeons des membres d'une souris polydactyle.

Expression du gène Shh dans le bourgeon du membre au jour 11,5 chez l'embryon de souris. B : bourgeon d'un membre avant d'un embryon de souris polydactyle. C : bourgeon d'un membre arrière d'un embryon de souris polydactyle.
Ces clichés concernant la souris polydactyle sont à comparer avec ceux de la souris normale. Outre l'expression de Shh dans la région postérieure du bourgeon, déjà présente chez un embryon normal, on trouve une expression dans la région antérieure du bourgeon aussi bien pour le membre avant que pour le membre arrière.
L’exploitation de ces documents permet donc d’établir une corrélation entre une extension du territoire d’expression du gène Shh au cours du développement et la polydactylie. Le fait qu’une protéine Shh non fonctionnelle empêche la formation des doigts, le fait que cette extension de l’expression dans la région antérieure a un effet comparable à la greffe d’une ZPA, indiquent que cette corrélation traduit une relation de causalité : c’est l’expression ectopique de Shh au cours du développement du membre qui engendre la polydactylie.
3 – La séquence ZRS régulatrice du gène Shh et l’extension de son territoire d’expression
Il s’agit maintenant d’établir ce qui cause l’expression ectopique du gène Shh dans la région antérieure du bourgeon du membre au cours du développement de l'embryon de souris polydactyle.
Chez la souris normale, on a mis en évidence une séquence, dite ZRS, qui contrôle l’expression du gène Shh dans le membre. Cette séquence est située à un million de paires de bases en amont du site de transcription de Shh. L'importance de la séquence non codante ZRS au cours du développement a été testée par des expérience où on a délété spécifiquement cette zone tout en gardant la séquence transcrite de Shh. Les figures suivantes permettent d'analyser les résultats de ces expériences.

En C et D, développement des embryons normal (à gauche) et de l'embryon chez lequel ZRS a été délétée à 14,5 jours de gestation. En F, souriceaux agés de 3 semaines.
Published by The Company of Biologists 2005.
On peut en déduire que la délétion de ZRS n'a pas d'effet sur le développement global de l'organisme (le souriceau est viable) mais entraîne l’absence de développement des doigts, la main étant réduite à un seul doigt. On a montré en outre que la délétion de ZRS entraîne l’absence d’expression de Shh au cours du développement du membre
On peut donc supposer que des mutations dans cette séquence ZRS sont impliquées dans la polydactylie. Les documents et données suivant vont permettre de tester cette hypothèse.
On a séquencé cette séquence ZRS chez les souris polydactyles et non polydactyles. La comparaison de ces séquences révèle une différence ponctuelle : la substitution d’une guanine (G) à une adénine (A) en position 488 (on a retenu une séquence de 540 pb).

Cette corrélation entre la différence de séquence de ZRS et la polydactylie, laisse à penser que c’est cette mutation ponctuelle de la séquence régulatrice qui entraîne l’expression ectopique de Shh. Cela a été testé par une expérience de transgénèse avec gène rapporteur.
Transgénèse avec gène rapporteur
Les chercheurs ont réalisé deux constructions génétiques. La première est constituée par la séquence ZRS normale associée au promoteur du gène Shh et à la séquence transcrite du gène rapporteur lacZ (voir gène rapporteur). La deuxième construction est identique à la première sauf que la séquence ZRS mutée remplace la séquence ZRS normale. Les chercheurs ont ensuite introduit à l’aide d’un micromanipulateur ces constructions dans des œufs de souris. Ils ont ensuite sacrifié les embryons à 11,5 jour de gestation. Ils ont réalisé des coupes dans ces embryons et les ont traitées avec la substance X-gal. L’enzyme beta galactosidase codée par le gène LacZ agit sur la substance X-gal avec formation d’un produit bleu. Les zones bleues de l’embryon indiquent donc les endroits où le gène rapporteur a été exprimé sous l’action de la séquence régulatrice ZRS. Ce sont bien sûr les endroits où le gène SHH se serait aussi exprimé sous l’action des séquences régulatrices ZRS.
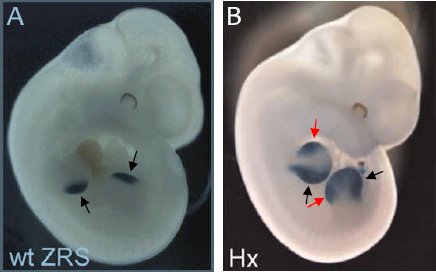
Expression de LacZ dans un embryon transgénique de 11,5 jours. A : sous contrôle de la séquence ZRS normale. B : sous le contrôle de la séquence ZRS mutée. Cette expression est observée sur un membre avant et un membre arrière. Les flèches noires indiquent la région postérieure des bourgeons des membres. Les flèches rouges la région antérieure.
hedgehog cis-regulator generate a variable regulatory out put responsible for preaxial
poydactyly” Human Molecular Genetics 17 (7):978-85.
On constate qu’avec la ZRS normale, l’expression du gène rapporteur est limitée à la région postérieure du bourgeon, alors qu’avec la ZRS mutée il y a en plus une expression dans la région antérieure. Autrement dit, les résultats de ces expériences de transgénèse miment les domaines d’expression de Shh dans les membres des embryons des souris normales et polydactyles. Cela confirme que la mutation ponctuelle dans la séquence régulatrice ZRS est à l’origine de la polydactylie.
4 - Retour à la polydactylie humaine
Familles avec polydactylie
|
|
Les symboles des personnes polydactyles sont en noir.
Les mécanismes génétiques causes de la polydactylie murine sont-ils saussi en jeu dans la polydactylie humaine ? Pour aborder cette question, on dispose des séquences du gène Shh et de la séquence ZRS chez les individus des arbres généalogiques 1 et 2.
La comparaison des séquences codantes des allèles du gène Shh montre qu’elles sont les mêmes chez les polydactyles et non polydactyles.
La comparaison des séquences ZRS révèle qu’il existe une différence ponctuelle qui n'est pas la même entre les personnes polydactyles et non polydactyles des deux familles. Les personnes polydactyles sont hétérozygotes pour la séquence ZRS ; on retrouve dans l’espèce humaine aussi les mêmes caractéristiques géniques trouvées chez la souris : pas de différence entre les séquences codantes du gène mais différence dans la séquence régulatrice de ce gène.
Bilan
L’analyse de l’origine du phénotype polydactyle montre donc comment un changement d’expression d’un gène de développement, une extension du territoire d’expression dans ce cas, entraîne un changement morphologique. Ce phénotype nouveau n’est pas dû à une mutation dans la séquence codante du gène mais à une mutation dans une séquence régulatrice du gène. On débouche donc sur un nouvel aperçu de la relation génotype-phénotype
Le gène de développement envisagé, Shh, intervient dans des régions différentes de l’organisme au cours du développement. La délétion du gène Shh entraîne des anomalies dans ces différentes régions, ce qui rend l’organisme non viable. En revanche, la délétion de la séquence régulatrice ZRS entraîne seulement des anomalies au niveau des membres et l’organisme est viable. Cela s’interprète en admettant qu’il existe plusieurs séquences régulatrices du gène Shh, chacune étant active dans une région bien définie (ZRS l'étant dans le membre).
Remarque : Bien entendu, cela n’est qu’une étape dans la compréhension des mécanismes génétiques en œuvre au cours du développement d’un membre. Il faut notamment expliquer pourquoi dans les conditions normales, l’expression de Shh est limitée à la région postérieure du bourgeon. Cela implique de connaître les facteurs de transcription qui régulent l’expression de Shh en se fixant à la séquence régulatrice. Les changements dans la séquence régulatrice de Shh modifient l’effet de ces facteurs de transcription. Ces facteurs de transcription étant codés par des gènes, c’est finalement l’idée de réseaux de gènes qui est sous-jacente. La protéine Shh elle-même n’est pas une protéine « effectrice » qui confère un phénotype particulier aux cellules où elle est produite. C’est une molécule de signalisation qui en agissant sur d’autres cellules va mettre en route le réseau génétique dont l’activité débouche sur un pattern de doigts bien défini.