
Exploitation pédagogique des facteurs de sélection
I - L’évolution de la pigmentation de la peau dans la lignée humaine
A - La répartition mondiale de la pigmentation
Le document sur la répartition mondiale de la couleur de la peau permet de dégager quelques grandes idées sur la variabilité de la couleur de la peau humaine et donc de sa pigmentation. Il est sous-entendu qu’il s’agit de la pigmentation constitutive de la peau, en dehors de toute exposition aux rayons solaires. On reconnaît une corrélation globale avec la latitude. Les peaux les plus sombres se trouvent à l’équateur et dans la zone intertropicale. Les peaux sont d’autant plus claires qu’on se dirige vers les latitudes élevées et cela est particulièrement net pour l’hémisphère nord, qui comprend plus de surfaces terrestres aux fortes latitudes que l’hémisphère sud. Le document sur la couleur des avant-bras montre qu’il n’y a toutefois pas de coupures nettes entre les couleurs des populations humaines mais un passage progressif des peaux les plus sombres aux couleurs les plus claires.
L’objectif est alors d’expliquer ces variations de la pigmentation en fonction de la latitude. L’important est de saisir que ce constat actuel de la diversité de la pigmentation est le résultat d’une évolution qui s’est déroulée durant l’histoire des populations d’Homo sapiens. Il est donc nécessaire d’avoir envisagé, au moins en partie, le thème « Un regard sur l’évolution de l’Homme » et d’avoir bien précisé l’origine africaine des Homo sapiens et leur dispersion hors de l’Afrique vers les différentes régions du monde et notamment les zones de forte latitude de l’hémisphère nord, à partir de 60000 ans environ. Les données sur les gènes de la pigmentation laissent à penser que les premières populations d’Homo sapiens avaient la peau sombre. Mais alors que les populations restées en Afrique ont conservé leur forte pigmentation, celles qui ont migré vers les latitudes élevées, ont subi une dépigmentation se traduisant par une peau claire. Il y a donc deux choses à expliquer : le maintien d’une forte pigmentation en Afrique et la dépigmentation des populations de l’hémisphère nord.
B - L’évolution de la couleur de la peau dans la lignée humaine, jusqu’aux premiers sapiens
Les premiers Homo sapiens africains avaient la peau fortement pigmentée. Il reste à voir comment cette couleur noire a été acquise par les Homo sapiens. Cela implique d’envisager la couleur de la peau chez les premiers représentants de la lignée humaine, les australopithèques et les premiers Homo erectus. On ne dispose pas d’indices fossiles renseignant directement sur la pigmentation de ces Hominines. Néanmoins, à partir de données phylogénétiques et physiologiques, on peut aboutir à un scénario plausible de l’évolution de la couleur de la peau dans la lignée humaine.
Il faut envisager en premier lieu la couleur de la peau de l’ancêtre commun à l’Homme et aux Chimpanzés. Même si cela est contre intuitif, la peau des Chimpanzés est assez peu pigmentée mais recouverte d’un abondant pelage noir. Cette couleur est apparente sur les régions glabres, notamment de la face du jeune Chimpanzé : cela représente le phototype constitutif de l’espèce. En vieillissant, la peau du visage, celle des mains, devient de plus en plus pigmentée par suite du bronzage sous l’action des UV du rayonnement solaire (pigmentation facultative). Mais sous les poils, la couleur de la peau reste claire. Ces caractéristiques de la peau se retrouvent chez les autres Primates catarrhiniens, à l’exclusion des Gorilles. On estime donc que c’est un état ancestral dans la lignée humaine et que l’ancêtre commun à l’Homme et au Chimpanzé devait aussi avoir une peau claire recouverte de poils sombres.

On ne dispose pas d’informations directes sur la peau des Australopithèques. On ne peut donc raisonner qu’à partir des caractéristiques anatomiques. Leur taille est voisine de celle des Chimpanzés, leur volume cérébral n’est que légèrement supérieur. Les caractéristiques de leur squelette indiquent qu’ils étaient bipèdes mais avaient aussi la capacité de grimper aux arbres. Leur bipédie était différente de celle des Homo et ne permettait pas de parcourir de longues distances. Bref, leur mode de vie n’était pas foncièrement différent de celui des Chimpanzés, ce qui fait qu’on estime que leur peau avait peu changé par rapport à celle de l’ancêtre commun : peau claire recouverte d’un pelage sombre.
|
|
Reconstruction of Australopithecus afarensis by Mauricio Antón | Reconstruction of early Homo erectus from Africa by Mauricio Antón. |


D'après N. Jablonski : Human Skin Pigmentation as an Example of Adaptive Evolution.
Les premiers représentants de l’espèce Homo erectus (au sens large) apparaissent en Afrique il y a 2 millions d’années. Par rapport aux Australopithèques, ils sont caractérisés par un squelette postcrânien très voisin de celui des Hommes modernes, et un plus grand volume cérébral. Leur bipédie exclusive leur confère la capacité de parcourir de longues distances, de courir. Ce sont des habitants de la savane actifs le jour. Pour ces Hommes très mobiles et actifs, il importe de pouvoir perdre la chaleur résultant de leur activité physique. C’est un problème, d’autant plus que ces Homo erectus africains vivent dans un environnement chaud. Disposer d’un mécanisme permettant d’évacuer la chaleur est une nécessité car une augmentation de quelques degrés de la température centrale (au-delà de 41-42°) peut entraîner la mort. La peau des Primates se caractérise, par rapport à celle de la plupart des autres mammifères, par la possession de glandes sudoripares sur toute la surface du corps. Celles-ci sont particulièrement abondantes chez les Hommes modernes par rapport aux autres Primates. Or l’évaporation de la sueur est un moyen de perdre de la chaleur dans un environnement chaud. Cela suggère qu’une évolution anatomique importante chez les Homo erectus a été l’augmentation de la densité des glandes sudoripares. Cependant, la possession d’un pelage dense diminue considérablement la perte de chaleur résultant de l’évaporation de la sueur. On estime que les individus au pelage peu dense régulaient mieux leur température centrale au cours de leurs activités physiques et avaient ainsi une plus forte probabilité d’arriver à l’âge adulte et donc de se reproduire. Ainsi sous l’action de la sélection naturelle, il y a eu une perte de la pilosité chez les Homo erectus africains. On ne sait exactement à quel moment elle a eu lieu, on estime généralement que le phénotype « peau nue » (à vrai dire à poils clairsemés et très fins) était acquis vers 1,2 millions d’années au moins et sans doute bien avant.
En conséquence les Homo erectus africains devaient avoir non seulement une peau nue mais aussi une peau peu pigmentée capable toutefois de bronzer. Or les données génétiques indiquent que les premiers sapiens africains avaient la peau noire (pigmentation constitutive). Au cours de l’évolution du genre Homo en Afrique, il y a donc eu un changement dans la couleur de la peau qui de claire est devenue noire.
En ce qui concerne l’évolution de la pigmentation de la peau, il faut donc envisager les facteurs qui ont conduit à l’acquisition d’une forte pigmentation de la peau chez les Homo africains et au maintien de cette pigmentation chez les populations africaines d’Homo sapiens. Bien entendu, il reste aussi à élucider les facteurs qui ont contribué à une dépigmentation de la peau des populations de sapiens qui ont quitté l’Afrique pour des régions de latitude plus forte.
II – Les données génétiques suggérant que la sélection naturelle est intervenue au cours de l’évolution de la pigmentation de la peau
Les études envisagées précédemment sur les gènes de la pigmentation (SLC24A5 ; MC1R et SLC45A2) ont révélé que ces gènes avaient un très faible polymorphisme dans les populations africaines avec une fréquence de près de 100% des allèles de ces gènes provoquant une forte pigmentation de la peau. Cela suggère qu’en Afrique dans les zones intertropicales et équatoriales, les allèles mutés pouvant entraîner une dépigmentation n’ont pas pu se répandre dans les populations et ont donc été éliminés par la sélection naturelle. En revanche pour le gène MC1R, les populations européennes et dans une certaine mesure asiatiques montrent un important polymorphisme avec une fréquence appréciable des allèles à l’origine d’une faible pigmentation. En ce qui concerne les gènes SLC24A5 et SLC45A2, la fréquence dans les populations européennes d’allèles mutés contribuant à une faible pigmentation est proche de 100%, donc de la fixation. Cela suggère que les contraintes qui conduisent à une élimination des allèles mutés « à peau claire » en Afrique sont levées aux latitudes élevées et que leur expansion dans les populations européennes peut être la conséquence d’une sélection naturelle positive.
Pour confirmer l’intervention de la sélection naturelle dans l’évolution et la différenciation de la couleur de la peau, il faut mettre en évidence les facteurs de sélection qui sont intervenus. Les populations humaines, lors de leur migration vers les latitudes élevées, ont rencontré des conditions d’environnement différentes de celles existant en Afrique. Parmi ces conditions, il faut identifier celles qui ont joué le rôle de facteurs de sélection.
L’étude de l’évolution de la pigmentation de la peau apparaît ainsi comme un excellent et motivant support pour traiter la partie du programme intitulée : « La biodiversité et sa modification » : Sous l’effet de la pression du milieu et du hasard, la diversité des populations change au cours des générations. L’évolution est la transformation des populations qui résulte de ces différences de survie et du nombre de descendants ».
III – L’intensité des UV et la pigmentation de la peau
Parmi les facteurs qui auraient pu avoir des répercussions sur la pigmentation de la peau humaine, l’attention des chercheurs s’est portée sur les variations spatiales de l’intensité du rayonnement solaire et plus particulièrement sur celles de sa composante UV.
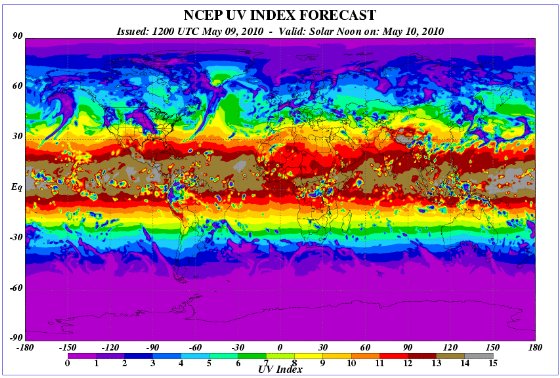
- Les documents (index-uv-mai-2010) et (index uv janvier 2014) permettent de comparer ponctuellement (au cours de deux journées, le 30 janvier et le 10 Mai) l’intensité du rayonnement UV entre les différentes régions du monde et à 2 moments de l’année. L’intensité de ce rayonnement est fournie par l’index UV dont la valeur varie de 0 à 15 ; cet index UV traduit l’intensité maximale du rayonnement UV le jour considéré (généralement entre 11h et 15h). Malgré le caractère ponctuel des données, on peut dégager les idées suivantes :
{kind=link}
{kind=link}
- Durant les deux jours, les intensités les plus fortes du rayonnement UV se trouvent dans les régions équatoriales et intertropicales ;
- L’intensité du rayonnement UV diminue lorsque la latitude augmente et cela aussi bien dans l’hémisphère nord que dans l’hémisphère sud.
Il semble exister des variations saisonnières de l’intensité des UV et cela est particulièrement net dans les régions de latitude élevée. Ainsi dans l’hémisphère nord, l’index 3-4 est atteint dès une latitude de 30° environ en janvier, alors qu'en Mai, cette valeur de l’index UV se trouve dans des régions de latitude supérieure à 60°.
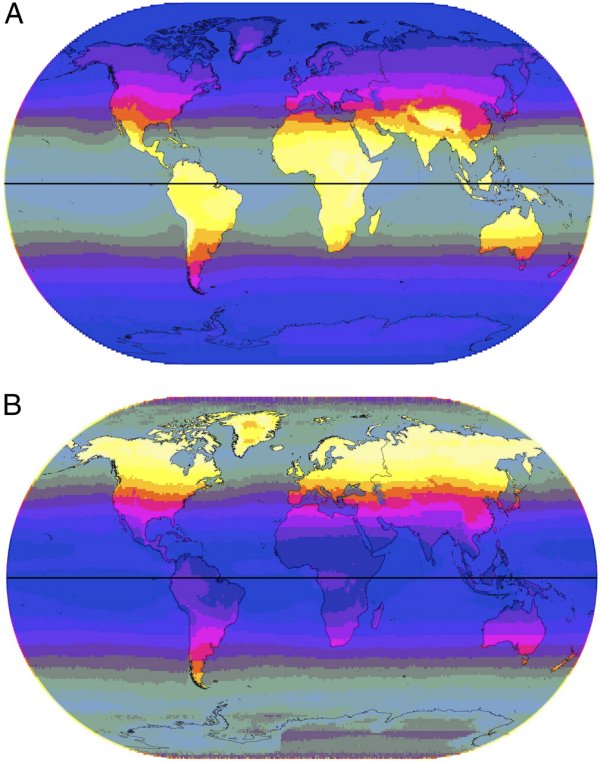
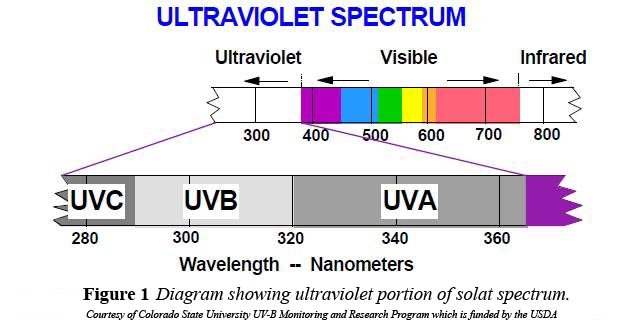
- Les documents UVB annuels et UVA annuels sont de portée plus globale car ils traduisent les moyennes annuelles de l’intensité du rayonnement UV aux différentes latitudes ainsi que l’importance des variations annuelles à chaque latitude. En outre, ils distinguent les rayonnements UVA et UVB. Les longueurs d’onde du rayonnement UV vont de 100 à 400nm (voir spectre UV) et on distingue les UVC (de 100 à 280nm), les UVB (de 280 à 320nm) et les UVA (de 320 à 400nm). Les UVC sont arrêtés par la couche d’ozone stratosphérique de sorte que seuls les UVA et UVB atteignent la surface de la terre et donc la peau humaine.
{kind=link}
{kind=link}
{kind=link}
De ces documents, on peut tirer les informations suivantes ;
- La légende indique que l’intensité du rayonnement UVB va de 1 à 135 Jm-2 alors que celle des UVA est comprise entre 65 et 930 Jm-2. Cela signifie que l’intensité des UVA est très nettement supérieure à celle des UVB ;
- Tant en ce qui concerne les UVB que les UVA, ce sont les régions équatoriales et intertropicales qui reçoivent le rayonnement le plus intense. Quasiment toute l’Afrique, une grande partie de l’Amérique centrale et du sud, ainsi que l’Australie sont exposées à un fort rayonnement UV ;
- L’intensité moyenne annuelle des UVB comme des UVA diminue avec la latitude ; les régions de forte latitude ayant une moyenne de rayonnement d’UVB très faible ;
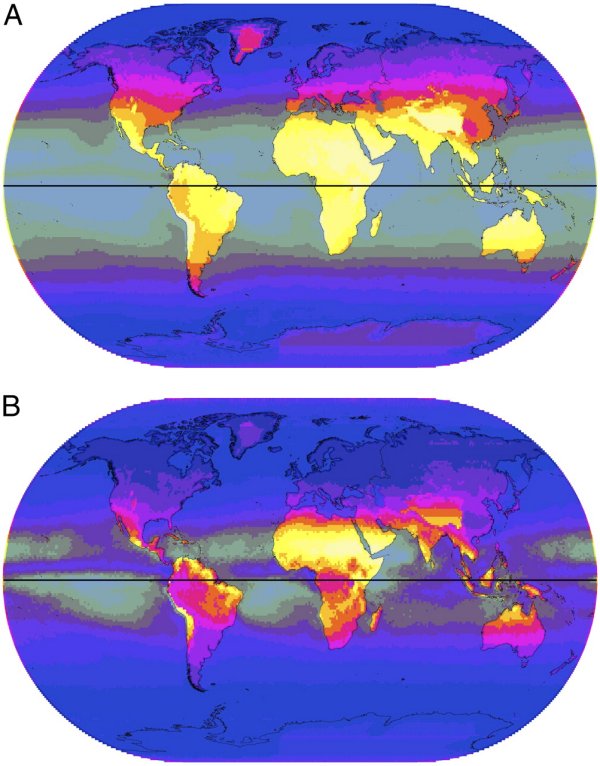
- La carte sur le coefficient de variation du rayonnement UVB au cours de l’année indique que c'est sont les régions situées au-delà de 50° nord environ que les variations des UVB sont les plus importantes. Cela traduit le fait que pendant plusieurs mois de l’année, les mois d’hiver, l’intensité des UVB est quasi nulle et qu’en été, tout en restant nettement inférieure à celle des régions intertropicales, elle est appréciable. La carte souligne que les régions intertropicales reçoivent toute l’année un rayonnement d’UVB assez uniforme (variations de l’ordre de 20%) ;
- Contrairement aux UVB, c'est sont dans les régions intertropicales que les variations de l’intensité des UVA sont les plus importantes, tout en restant nettement moins fortes que celles des UVB. Cela est dû que même en hiver, les régions de forte latitude reçoivent un rayonnement d’UVA conséquent et qu’il n’augmente pas considérablement durant l’été.
En résumé, le fait le plus remarquable des variations spatiales de l’intensité des UVB et des UVA est la corrélation qui existe avec la latitude. Or la pigmentation est aussi corrélée avec la latitude. On peut donc en déduire qu’il y a une corrélation entre l’intensité du rayonnement UV et la pigmentation de la peau : une peau sombre fortement pigmentée dans les régions de rayonnement UV intense et assez uniforme toute l’année ; une peau claire, peu pigmentée là où le rayonnement d’UV et particulièrement celui des UVB est faible ; une peau de couleur intermédiaire dans les régions d’intensité UV moyennes.
Les premiers Homo sapiens africains avaient une peau noire, héritage des Homo dont ils descendent ; dans l’histoire de la lignée humaine, cette peau noire était un état dérivé. Chez les populations de sapiens qui ont quitté l’Afrique, il y a 60000 ans environ pour peupler l’Eurasie, zones où l’intensité du rayonnement UV, en particulier UVB, est nettement plus faible, il y a eu une dépigmentation. En revanche les populations de sapiens qui sont restées en Afrique, à fort rayonnement UV, ont conservé leur peau sombre.
Ces données suggèrent que ce sont les différences dans l’intensité et les caractéristiques du rayonnement UV auquel ont été soumises les populations humaines au cours de leur histoire qui sont cause des variations de couleur de la peau constatées aujourd’hui. Pour corroborer cette hypothèse, il faut envisager comment le rayonnement UV peut influencer la pigmentation constitutive de la peau.
IV – La sélection naturelle et l’acquisition, le maintien d’une forte pigmentation dans les régions où le rayonnement UV est intense
Les données génétiques suggèrent que la sélection naturelle a joué un rôle majeur dans l’acquisition d’une peau fortement pigmentée chez les Homo africains. Avant d’aborder l’exploitation des documents (Les UV et l’ADN et les UV et les folates) qui permettent de tester cette hypothèse, il est sans doute utile de rappeler les principes de la sélection naturelle déjà envisagés en classe de seconde.
- Le premier point est l’existence d’une variabilité phénotypique au sein des populations : dans le cas des populations d’Homo africains, l’apparition de variants à peau fortement pigmentée au sein de populations à la peau claire.
- Le deuxième point est que cette variabilité phénotypique ait un déterminisme génétique et soit donc transmissible.
- Le troisième point est que cette diversité phénotypique dans les conditions d’environnement s’accompagne d’un succès reproducteur différentiel : les personnes à peau noire devraient contribuer davantage à la génération suivante que les personnes à peau claire. L’étude des gènes de la pigmentation de la peau faite précédemment et des mutations que peuvent subir ces gènes, montre que les deux premiers points sont satisfaits. Il reste à voir si un environnement avec un rayonnement UV intense peut avoir un impact sur un succès reproducteur différentiel entre les individus à peau claire et ceux à peau fortement pigmentée.
En classe de première, on a vu que les UV sont des agents mutagènes. Les documents sur « les UV et l’ADN » permettent d’aborder le mécanisme en jeu et d’en voir les conséquences. Le spectre d’absorption de l’ADN indique que l’ADN absorbe les UVB et le spectre d’action souligne que cette absorption entraîne des dommages à L’ADN. L’ADN n’absorbe pas les UVA mais le spectre d’action souligne que malgré tout les UVA, en particulier ceux de longueur d’onde comprise entre 320 et 340nm causent aussi des dommages à l’ADN. Le texte indique pourquoi : ils provoquent la formation dans les cellules de radicaux oxygénés qui sont agressifs vis-à-vis de l’ADN. Les figures soulignent la nature des dommages causés à L’ADN , notamment la distorsion de la molécule d’ADN causée par la formation de dimères sous l’action directe ou indirecte des UV. Le point important est que ces dommages sont à l’origine des mutations pouvant engendrer des cellules cancéreuses.
Une manière d’aborder le document sur « la pigmentation de la peau et les dommages à l’ADN » est de comparer l’incidence des cancers cutanés chez les personnes à peau noire et celles à peau claire. Elle est beaucoup plus faible chez les personnes à peau fortement pigmentée ce qui suggère que la mélanine beaucoup plus abondante chez les personnes noires protège contre les dommages à l’ADN causés par les UV. Le spectre d’absorption de la mélanine indique que ce pigment absorbe les radiations du spectre solaire depuis les UVB jusqu’aux radiations infra rouges. La richesse en mélanine de l’épiderme des personnes à peau noire fait que qu’il forme un écran protecteur et qu’en conséquence l’intensité des UV atteignant le derme soit 5 fois plus faible chez les noirs que chez les blancs. Le document sur la pénétration des UV dans la peau illustre ce rôle protecteur des couches épidermiques supérieures riches en mélanine notamment dans le blocage des UVB.
Considérons alors les populations d’Homo erectus initialement à peau claire chez lesquelles apparaissent suite à des mutations des individus ayant la peau plus foncée, riche en mélanine. Ceux-ci ont une probabilité plus faible que le reste de la population d’être victime d’un cancer cutané. Peut-on dire pour autant que cela a contribué à l’évolution de la coloration de la peau de ces populations d’Homo jusqu’à ce qu’elle soit entièrement constituée par des personnes à peau noire ?
Le document sur l’incidence des cancers cutanés en fonction de l’âge permet d’aborder cette question de façon critique On voit que tant ce qui concerne les carcinomes que les mélanomes, la fréquence de ces cancers ne devient appréciable qu’à partir de 45-50 ans, c'est-à-dire bien après l’atteinte de la maturité sexuelle. En outre l’espérance de vie devait être nettement inférieure à celle d’aujourd’hui. Il est donc fort peu probable que la protection assurée contre l’apparition de cancers cutanés par la mélanine ait joué un avantage sélectif chez les personnes à peau sombre par rapport à celles à peau claire. Les dommages causés à l’ADN par les UV ne sont pas ou très peu impliqués dans l’évolution de la pigmentation des populations d’Homo africains. Cet exemple est excellent pour bien faire saisir ce qu’est la sélection naturelle. Des différences de survie entre phénotypes différents n’entraînent une évolution dans les populations que dans la mesure où elles entraînent une reproduction différentielle de ces phénotypes. La plus grande fréquence des cancers cutanés chez les individus à peau claire n’affectait pas leur capacité de transmette leurs allèles des gènes de la pigmentation à la génération suivante.
Avec les documents sur « Les UV et les folates », on aborde un autre aspect de l’action des UV sur l’organisme. Le premier document de cette partie permet de sensibiliser à l’importance des folates. Le graphique indique qu’en l’absence de folate dans le milieu de culture des kératinocytes, il n’y a pas de croissance de la population cellulaire en culture. L’ajout de folate au milieu entraîne un accroissement de la population cellulaire et cela d’autant plus qu’il est important. Les folates sont donc des nutriments indispensables à l’accomplissement des cycles cellulaires. Cela se comprend car ils sont indispensables à la synthèse des nucléotides. En conséquence une déficience en folates va avoir un fort impact aux périodes du développement où le rythme des mitoses est élevé, notamment durant la phase embryonnaire. Cela est à l’origine des défauts dans la construction du système nerveux en cas de déficience en folates de la mère. La déficience en folates provoque aussi une défaillance de la spermatogenèse, l’efficacité de celle-ci dépendant du rythme des mitoses des spermatogonies. Le document aborde l’action des UV sur la concentration plasmatique des folates. On constate d’abord que l’acide folique absorbe les UVB et à un moindre degré les UVA. L’expérience de Branda et Eaton montre que lorsqu’on expose du plasma aux rayons UV pendant 60 minutes cela entraîne une diminution de la concentration plasmatique en folates de 30à 50%.Cela indique que l’absorption du rayonnement UV par les folates provoque leur dégradation comme le confirme le document sur l’acide folique. La comparaison des deux groupes de personnes, le premier exposé à des UV chaque jour pendant trois mois, le second non exposé jouant le rôle de témoin indique que l’exposition prolongée aux UV entraîne une baisse de la concentration du plasma en folates de 7 à 4ng/ml. Cela est confirmé par les données de Shadeen.
Il reste à préciser comment les UV entraînent une chute de la concentration en folates du plasma. L’épiderme n’est pas vascularisé et c’est en circulant dans les capillaires dermiques que les folates du plasma sont dégradés. En effet, et en particulier chez les personnes à peau claire,, les UVA arrivent jusqu’au derme et peuvent donc entraîner une photolyse des folates. Cette dégradation est beaucoup plus minime chez les personnes à peau sombre à cause de l’écran constitué par la richesse en mélanine qui bloque la majorité des UVA.
Revenons aux populations africaines d’Homo soumises à un rayonnement UV intense. Les UV provoquaient une déficience en folates beaucoup plus forte chez les personnes à peau claire que chez les personnes à peau sombre. Or cette déficience retentit sur le développement, la spermatogenèse, dont affecte la capacité de l’individu à contribuer à la genèse de la génération suivante. Le phénotype peau claire dans ces conditions d’environnement avait un désavantage sélectif par rapport au phénotype peau sombre. Ainsi de génération en génération, la fréquence du phénotype peau fortement pigmentée est devenue de plus en plus grande jusqu’à ce qu’il soit fixé dans la population. La fréquence des allèles des gènes de la pigmentation contribuant à une forte mélanisation de la peau a atteint une fréquence proche de 100%. Et ce très faible polymorphisme s’est prolongé au cours de l’histoire des sapiens africains car les allèles à l’origine d’une dépigmentation ont été éliminés par la sélection naturelle (sélection négative, « purificatrice ».
V - La sélection naturelle et la dépigmentation de la peau au cours de l’évolution des populations européennes et asiatiques d’Homo sapiens
Les populations d’Homo sapiens qui ont quitté l’Afrique pour les régions de plus forte latitude de l’hémisphère nord ont vécu dans des environnements où le rayonnement UV et notamment celui des UVB est nettement moins intense que dans les régions intertropicales et équatoriales. Au cours de la colonisation de ces régions, les populations ont subi une dépigmentation de la peau, conduisant aux peaux claires des populations asiatiques et européennes Pour résoudre le mécanisme de cette dépigmentation , on dispose de documents qui permettent d’établir qu’en dehors de leurs effets nocifs, les UV, plus précisément les UVB ont un effet bénéfique car contribuant à la synthèse de vitamine D. Les variations de l’impact des UVB sur la synthèse de vitamine D en fonction de différents facteurs fournissent les données qui permettent d’expliquer la dépigmentation de la peau chez les populations européennes et asiatiques.
Données essentielles sur la biologie de la vitamine D
Le graphique sur l’évolution de la concentration de la vitamine D suite à l’exposition à un rayonnement solaire artificiel, indique une augmentation de la concentration plasmatique de cette vitamine de plus de 150% au bout de 5 semaines. Sous l’effet du rayonnement solaire, l’organisme produit donc de la vitamine D.
La figure sur la biologie de la vitamine D précise que c’est au niveau de la peau qu’est synthétisée la vitamine D, plus précisément la vitamine D3 qui est ensuite transformée par le foie et le rein en vitamine D définitive. Il y a donc synthèse de vitamine D3 sous l’action du rayonnement solaire. Le point crucial, mentionné dans le texte est que ce sont les UVB qui assurent cette synthèse. Ce sont les cellules épidermiques de la couche basale et celles qui la surmontent qui sont le siège de la synthèse de vitamine D3. Elles possèdent dans leurs membranes plasmiques un précurseur, le 7 déhydrocholestérol, qui absorbe les UVB, suite à quoi, il se transforme en vitamine D3. Là encore c’est une réaction photochimique qui est en jeu. La vitamine D3 fabriquée par les cellules épidermiques, diffuse dans le derme et passe dans le sang du réseau de capillaires dermiques. Le reste du texte permet d’apercevoir l’importance physiologique de la vitamine D.
Une forte déficience en vitamine D peut avoir des conséquences sur la survie d’ l’individu et cela dès le jeune âge. Par-là elle peut diminuer la capacité d’une personne à contribuer à la genèse de la génération suivante.
Je pense qu’il faut modifier l’ordre des documents retenu jeudi dernier.
La pigmentation de la peau et la synthèse de vitamine D3 par la peau
L’expérience in vitro montre que sous l’effet du rayonnement solaire, il y a synthèse de vitamine D3 par le fragment cutané à peau claire et que cette synthèse augmente avec le temps d’exposition. En revanche, pour le fragment à peau sombre soumis au même rayonnement solaire, la synthèse de D3 est beaucoup plus faible et n’augmente pas au-delà de 10 minutes d’exposition. On en conclut qu’une forte pigmentation de la peau diminue la capacité de celle-ci à produire de la vitamine D sous l’action des UVB. A partir des connaissances précédemment acquises sur la mélanine, on peut proposer une explication : la mélanine ayant un large spectre d’absorption et absorbant notamment les UVB, elle diminue l’intensité des UVB arrivant aux cellules épidermiques productrices de vitamine D3. Le rôle d’écran protecteur vis-à-vis des effets nocifs des UV s’avère défavorable à la synthèse de vitamine D3.
Le graphique relatif aux effets in vivo de la pigmentation de la peau sur la synthèse de vitamine D3 confirme les conclusions extraites de l’expérimentation in vitro et les complète.
L’exposition aux UVB entraîne chez les trois personnes à peau claire une variable mais toujours nette augmentation de la concentration plasmatique de D traduisant une synthèse cutanée de vitamine D3. La même exposition ne provoque quasiment pas d’augmentation de la concentration de vitamine D3 chez les personnes à peau sombre. La richesse en mélanine freine donc la production cutanée de D3. Toutefois si l’intensité du rayonnement UV est 6 fois plus intense, la concentration en vitamine D3 augmente nettement sans atteindre toutefois les valeurs des personnes à peau claire. Les peaux fortement pigmentées sont donc capables de produire de la vitamine D3 mais pour produire la même quantité de D3 que les personnes à peau claire il leur faut un rayonnement UVB plus de 5 fois plus intense.
La pigmentation de la peau et la synthèse de vitamine D
Là il faut modifier le graphique fourni. De façon incompréhensible (il s’agit des mêmes auteurs) le document retenu extrait de « factors that » n’a pas les mêmes ordonnées que celui présent dans « photobiology of vitamin D). Je préfère ce dernier ‘plus en accord avec un document précédent.
Le document sur la synthèse de vitamine D en fonction du phototype,, permet de voir si les conclusions dégagées à propos de la synthèse de vitamine D3 se retrouvent lorsqu’on considère la vitamine D définitive. On constate d’abord que pour tous les types de peau, il y augmentation de la concentration en vitamine D suite à l’exposition au rayonnement solaire : l’augmentation de la production cutanée de vitamine D3 se traduit bien par une plus forte concentration plasmatique de la vitamine D. En outre, cette augmentation de la concentration de vitamine D sous l’action du rayonnement solaire est d’autant plus importante que la peau est claire ; en effet les pourcentages d’augmentation pour les phototypes 11 :111 ; 1V et V sont respectivement de :210 ;190 ;125 et 40%
Influence de la latitude et des saisons sur la production cutanée de vitamine D3
En regardant attentivement, on s’aperçoit que le graphique n’est pas net toujours net, notamment pour Edmonton : il faut indiquer qu’on ne dispose pas de données pour cette localité de ? ou alors supprimer la courbe relative à Edmonton si cela est possible
Le graphe A permet d’étudier l’influence des saisons et de la latitude sur la production de vitamine D3 par une personne à la peau claire dans trois localités de l’hémisphère nord. Elle est négligeable durant les mois de décembre à février à Boston (42°N), de novembre à mars à Edmonton (52°N) et d’octobre à Mars à Bergen ‘61°N.La production de vitamine D3 s’interrompt donc dans ces localités durant les mois d’hiver et ce d’autant plus longtemps que la latitude est élevée. On parle « d’hiver de la vitamine D » ; En outre l’intensité de la production de vitamine D durant les mois d’été est plus importante à Boston que dans les deux autres localités ; non seulement la durée annuelle de production de la vitamine D mais aussi son intensité diminue avec la latitude.
On peut en fournir une explication en se rapportant aux variations annuelles des UVB précédemment envisagées. : l’intensité des UVB diminue avec la latitude et pendant les mois d’hiver l’intensité des UVB est très faible aux fortes latitudes.
La comparaison des graphes A et C permet de retrouver l’effet de la pigmentation de la peau sur la pigmentation de la peau. A Boston par exemple , la période « d’hiver de la vitamine D »s’étend d’octobre à mars pour les personnes à la peau sombre soit trois mois de plus que pour les personnes à peau claire.
Les données relatives à l’hémisphère sud confirment celles de l’hémisphère nord : la production de vitamine D diminue avec la latitude. On peut surtout noter que pour la localité située à la plus faible latitude, Johannesburg (26°S), il y a une production cutanée de vitamine D pendant tous les mois de l’année, la synthèse de D3 pendant les mois d’hiver étant encore appréciable. Cela est vrai même pour les personnes à peau sombre. Cela doit être aussi le cas dans les régions intertropicales et équatoriales de latitude plus faible (‘tropique du capricorne :23° environ). ; Autrement dit les personnes à peau noire n’ont pas de problème pour synthétiser constamment de la vitamine D dans les régions intertropicales malgré la richesse en mélanine de leur peau et ce à cause du fort rayonnement UVB durant toute l’année.
La sélection naturelle et l’évolution de la couleur de la peau
La figure ci-dessous résume deux effets majeurs de l’action des UVB sur la peau ayant eu un fort impact sur l’évolution de sa coloration. Mettre là la figure extraite de l’article de Jablonski 2004)
Dans les régions intertropicales et équatoriales à fort rayonnement UV pendant toute l’année, la richesse en mélanine protège contre les effets destructeurs des folates ; cependant le rayonnement UV est suffisamment intense pour permettre une synthèse suffisante de vitamine D pendant toute l’année. Les individus à peau peu pigmentée peuvent aussi synthétiser toute la vitamine D nécessaire (il n’y a pas de risque d’hypervitaminose D car le rayonnement ultraviolet détruit la vitamine D3 si elle est trop abondante) mais leur faible pigmentation ne les protège pas contre la destruction des folates. Il en résulte une pression sélective négative des individus à peau claire par rapport à ceux à peau sombre ce qui explique l’acquisition d’une peau fortement pigmentée chez les Homo et le maintien de cette pigmentation chez les sapiens africains.
Au cours de la migration des sapiens à peau noire vers l’Eurasie, il y a eu des mutations des gènes de la pigmentation qui ont eu pour effet l’apparition d’individus à peau plus claire. Une variation phénotypique de la couleur de la peau est apparue dans ces populations. Dans ces régions de l’Eurasie de latitude moyenne ou forte, ces populations ont été soumises à un rayonnement UV nettement moins intense et très variable annuellement surtout pour les UVB. La forte pigmentation des individus à peau noire ne leur permettait pas de synthétiser la vitamine D pendant quelques mois et annuellement leur production de vitamine D était déficiente. Les individus à peau claire du fait de la réduction du rayonnement UV risquaient beaucoup moins une destruction de leurs folates et grâce à leur faible pigmentation produisaient suffisamment de vitamine D (cette vitamine fabriquée majoritairement en été peut être stockée transitoirement dans le foie). En conséquence dans ces populations migrantes de sapiens, les individus à peau noire ont été soumis à une pression sélective négative et le phototype peau claire s’est généralisé. Les gènes SLC24A5 et SLC45A2 où les allèles mutés entraînant une faible pigmentation ont une fréquence de presque de 100% dans les populations européennes traduisent ce processus de sélection. Il faut remarquer que ces allèles ne se retrouvent pas dans les populations asiatiques qui ont pourtant une peau claire. Cela laisse à penser que les mutations à l’origine de la faible pigmentation des européens ont eu lieu après la séparation des populations de sapiens européens et asiatiques. En outre cela implique que les innovations génétiques à l’origine de la dépigmentation des européens et asiatiques sont différentes bien que se traduisant par un phénotype semblable (évolution convergente).