
Exploitation pédagogique Souris Agouti
Note : Première S
1 - Effet de deux gènes sur la couleur du pelage de la souris
- Dans l’étude des rôles du génotype et de l’environnement dans la réalisation d’un phénotype, l’exemple de la couleur du pelage de la souris permet d’envisager en premier l’interaction de deux gènes, le gène MC1R et le gène agouti dans la réalisation de ce caractère.
- Pour le gène MC1R, il faut consulter les documents : « MC1R et la couleur du pelage de la souris&nbsnbsp;» et « La protéine codée par le gène MC1R » ainsi que le début de l’exploitation pédagogique relative à MC1R. On envisage les phénotypes de deux mutants, l’un à pelage jaune, l’autre à pelage noir. Ces phénotypes sont dus à deux allèles mutés du gène MC1R : l’un code pour une protéine non fonctionnelle et est à l’origine du phénotype « pelage jaune » de la souris ; l’autre code pour une protéine dont l’activité est plus forte que celle codée par l’allèle « sauvage » et est à l’origine du phénotype « couleur noire du pelage ».
- Le document : « Le gène agouti et la couleur du pelage de la souris » porte aussi sur le déterminisme génétique du phénotype « pelage noir ». Les résultats des croisements réalisés indiquent que ces souris noires diffèrent des souris « sauvages » par un seul des gènes intervenant dans la couleur du pelage. L’étude précédente sur le gène MC1R laisse à penser que ces souris noires possèdent un allèle muté du gène MC1R. La comparaison avec Anagène, des séquences des allèles MC1R d’une souris grise et de la souris noire envisagée révèle que ces allèles sont identiques. Autrement dit, le phénotype « pelage noir » est dû à un allèle d’un autre gène. Ce gène a été isolé et dénommé le gène agouti (on désigne souvent le phénotype sauvage - pelage brun ou gris par phénotype agouti).
- La comparaison des séquences codantes des allèles du gène agouti chez la souris sauvage (phénotype agouti) et chez la souris noire mutante indique que l’allèle muté diffère par une délétion de 3 nucléotides CTC au codon 49, suivie d’une insertion de 2 nucléotides TA. Il en résulte un décalage du cadre de lecture et une séquence modifiée aux codons 49-54. Le codon 55 de l’allèle muté devient un codon stop. La protéine codée par cet allèle ne comprend que 54 acides aminés au lieu de 131, ce qui la rend non fonctionnelle.
- Il faut alors exploiter les données sur les protéines MC1R et agouti pour expliquer comment des mutations du gène MC1R et du gène agouti conduisent au même phénotype : « pelage noir ».
- La protéine MC1R code pour une protéine membranaire exprimée à la surface des mélanocytes. Lorsque cette protéine est non fonctionnelle, le phénotype de pelage est jaune ce qui indique une synthèse de phéomélanine par le mélanocyte indépendante de MC1R. La protéine MC1R « sauvage » entraîne donc une synthèse d’eumélanine par le mélanocyte qui synthétise les deux types de mélanine, surtout de l’eumélanine, à l’origine du phénotype agouti (couleur brune). Si la protéine MC1R est plus active que la protéine sauvage, alors la synthèse d’eumélanine est plus forte, celle de phéomélanine quasi inexistante, et le phénotype est « pelage noir ».
- La protéine MC1R est un récepteur membranaire. Il a une activité de base qui peut être modifiée par les messagers qui s'y lient. Ces messagers sont l’hormone MsH qui le stimule à produire de l’eumélanine, et la protéine agouti sécrétée par des cellules du derme, qui l’inhibe. Lors de la croissance des poils, les deux messagers agissent sur les mélanocytes par l’intermédiaire des récepteurs MC1R. Si la protéine agouti est non fonctionnelle, la synthèse d’eumélanine n’est plus inhibée et les mélanocytes ne synthétisent plus que de l’eumélanine d’où le phénotype « pelage noir ».
Le schéma ci-dessous illustre comment les interactions entre les deux gènes MC1R et Agouti contrôlent la couleur du pelage.
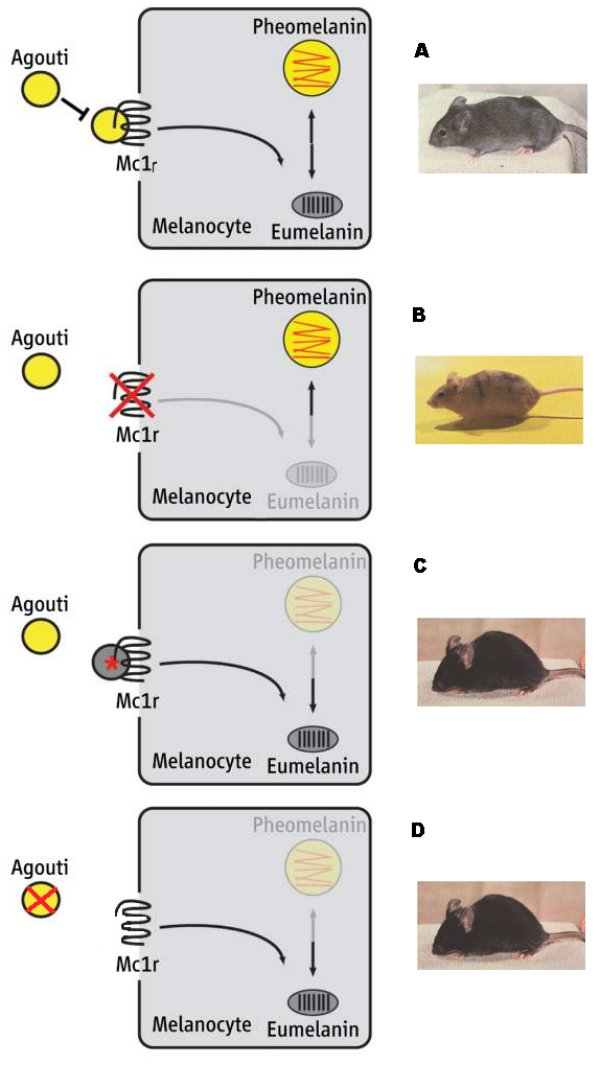
A - Protéine agouti et récepteur MC1R fonctionnels : synthèse de phéomélanine et d'eumélanine, d'où la couleur grise (phénotype agouti).
B - Protéine agouti fonctionnelle, récepteur MC1R non fonctionnel : synthèse uniquement de phéomélanine, d'où la couleur jaune.
C - Protéine agouti fonctionnelle, récepteur MC1R hyperactif : synthèse uniquement d'eumélanine, d'où la couleur noire.
D - Protéine agouti non fonctionnelle, récepteur MC1R fonctionnel : synthèse uniquement d'eumélanine, d'où la couleur noire.
2 – Effets des variations de l’expression d’un gène sur la couleur du pelage de la souris
- Le document sur « l’allèle Avy du gène agouti et le phénotype souris jaune », montre une souris obèse au pelage jaune à côté d’une souris agouti au phénotype sauvage. Dans le document « le gène MC1R et la couleur pelage de la souris », on a déjà vu que le phénotype pelage jaune pouvait être dû à une mutation du gène MC1R rendant non fonctionnel ce récepteur (sans obésité). L’obésité de cette souris laisse à penser que le gène MC1R n’est pas en cause ce que confirme le texte sur l'allèle Avy. On pense alors à une mutation du gène agouti. La comparaison des séquences codantes des allèles du gène agouti chez la souris agouti et la souris jaune ne révèle cependant aucune différence. L’information relative à la synthèse d’ARNm du gène agouti souligne que l’allèle de la souris jaune est surexprimé par rapport à l’allèle sauvage. Chez la souris jaune, la surexpression du gène agouti entraîne une forte production de la protéine agouti qui inhibe la protéine MC1R et bloque donc la synthèse d’eumélanine au bénéfice de celle de phéomélanine, d’où la couleur jaune du pelage.
Le schéma ci-dessous illustre le rôle des deux gènes dans la réalisation du phénotype :
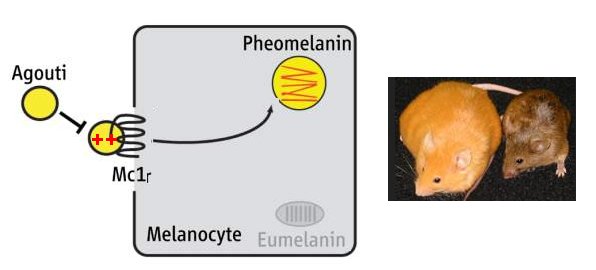
Protéine agouti et récepteur MC1R fonctionnels ; sur expression de l'allèle Avy d'où forte production de la protéine agouti qui inhibe le récepteur MC1R, ce qui empêche la production d'eumélanine et explique la couleur jaune. Noter un autre caractère phénotypique : l'obésité.
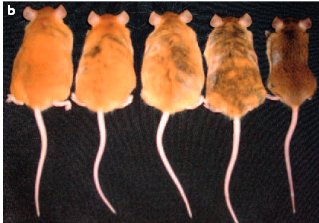
- La sur expression des allèles du gène agouti chez les souris jaunes est due à l’insertion d’une séquence d’ADN, nommée IAP dans la région régulatrice du gène agouti.
- En conclusion de ces deux premiers points, on voit que le phénotype de la couleur du pelage peut varier à la suite d'une mutation de la séquence codante d’un gène qui modifie l’activité de la protéine qu’il code, mais aussi à la suite d’une mutation qui affecte la séquence régulatrice du gène et peut en modifier l’expression (intensité de son expression et, comme on verra, territoire de l’expression). L’allèle du gène agouti responsable de la couleur jaune du pelage est dénommé Avy (Agouti viable yellow).
3 - Un même génotype et des phénotypes différents
- L’exploitation de ce document nécessite de réinvestir la connaissance des caractéristiques des allèles a et Avy du gène agouti. Un simple échiquier de croisement indique que les génotypes de la génération F1 issue du croisement sont : 50% a//a et 50% Avy//a. Comme la protéine codée par l’allèle a est non fonctionnelle, le pelage de ces souris a//a est noir ce qui est en accord avec les données fournies (50% de souris noires). Les F1 de génotype Avy //a devraient avoir le même phénotype ; or on constate que ce n’est pas le cas. Le problème à résoudre est donc le suivant : comment des souris ayant le même génotype peuvent-elles avoir des phénotypes différents ?
- Puisque l’allèle a est non fonctionnel, le phénotype des F1 de génotype Avy//a dépend de l’expression de l’allèle Avy. Considérons alors les deux phénotypes extrêmes : pelage jaune et pelage agouti (brun). Le pelage jaune s’explique par une forte expression de l’allèle Avy qui fait que les mélanocytes des follicules pileux synthétisent uniquement de la phéomélanine. Le phénotype pseudo agouti est semblable au phénotype sauvage. Il implique que l’allèle Avy s’exprime comme l’allèle sauvage du gène agouti, ce qui conduit à la synthèse surtout d’eumélanine mais aussi de phéomélanine par les mélanocytes des follicules pileux. Les deux phénotypes « jaune » et « pseudo agouti » sont donc dus à des expressions différentes de l’allèle Avy suivant les individus et non à des différences dans la protéine qu’il code : sur expression de l'allèle dans un cas, expression normale dans l’autre cas. Les autres phénotypes apparaissent comme des intermédiaires entre les deux phénotypes extrêmes, se rapprochant soit du phénotype « pelage jaune », soit du phénotype « pelage brun ». On peut donc penser que l’expression de l’allèle Avy est décroissante du phénotype pelage jaune au phénotype pelage brun en passant par les phénotypes « légèrement tacheté » « tacheté » « très tacheté ». A vrai dire, chez les souris au phénotype tacheté coexistent des endroits où le pelage est jaune (forte expression de l’allèle Avy et d’autres (taches) où le pelage est brun (expression de base de l’allèle Avy).
- Le problème à résoudre est alors celui du mécanisme à l’origine de cette variabilité dans l’expression de l’allèle Avy suivant les individus étant donné que des différences entre d’autres gènes impliqués dans la pigmentation ne semblent pas intervenir.
- Le document Un même génotype et des phénotypes différents présente des modifications qui peuvent affecter l’ADN d’un gène en particulier dans les zones régulatrices (au sens général, comprenant le promoteur) sans modifier pour autant la séquence des nucléotides du gène. Une modification particulièrement fréquente est la méthylation de l’ADN qui a un effet inhibiteur sur la transcription d'un gène.
- Jirtle et Waterland, chercheurs qui ont travaillé sur le modèle de la souris agouti, ont cherché à savoir si la variabilité de l’expression de l’allèle Avy suivant les individus pouvait être due à des variations dans la méthylation de cet allèle et plus précisément du segment d’ADN inséré dans la région régulatrice (IAP - intracisternal A-particle), segment qui fait l’originalité de cet allèle.
- On dispose de la séquence de ce segment. On peut demander aux élèves de rechercher les sites de cette séquence qui pourraient être méthylés et de s’apercevoir qu’ils sont nombreux.
GGCACCAATTGTTATTCGACGCGTTCTCACGACCGGCCAGGAAGAACACCACAGACCAGAATCTTC
TGCGACAAAGCTTTATTCTTACATCTTCAGGAAAAGAGAGCAAGAAGCAAGAGAGAGAGAAAAACGAA
ACCCCTTCTATTTTAAAGAGAACAACCATTGCCTAGGGCGCATCACTCCCTGATTGGCTGCAGCCCAT
GGCCGAGCTGACGTTCACGGGAAAAACAGAGTACAAGTGGTCGTAAATACCCTTGGCTCATGCGCAG
ATTATTTGTTTACCAACTTAGAACACAGGATGTCAGCGCCATCTTGTGACGGCGAATGTGGGGGCGGC
TTCCCACA
Ce segment IAP est un ilôt de sites propices à la méthylation.
- L’histogramme dans le document indique que le pourcentage de méthylation de Avy très faible (environ 5%) chez les souris jaunes, augmente d'autant plus que la surface de la peau de couleur brune est grande pour atteindre une valeur maximale chez les souris de phénotype brun (pseudo agouti). Il existe donc une corrélation inverse entre la méthylation de l’allèle Avy et l’intensité de son expression. : plus il est méthylé moins l’intensité de son expression est forte. Etant donné ce qu’on sait par ailleurs de l’effet de la méthylation, cette corrélation traduit une relation de cause à effet.
Les figures qui suivent schématisent les relations entre le degré de méthylation de l'allèle Avy et le phénotype couleur du pelage.
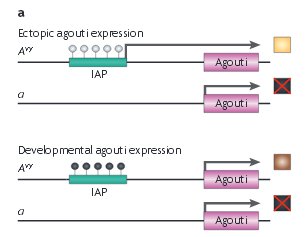
En blanc les sites non méthylés, en noir les sites méthylés. On n'a représenté que les méthylations des deux types extrêmes. Pour les autres phénotypes la méthylation est intermédiaire. Les flèches indiquent le phénotype cutané dû à l'allèle Avy.
D'après Environmental epigenomics and disease susceptibility. Jirtel et Skinner.
D'après Get the Lead Out of Your Epigenome
- On qualifie d’épigénétiques les mécanismes qui agissent sur l’expression d’un gène sans modifier la séquence de nucléotides de ce gène. La méthylation de l’ADN est un de ces mécanismes. Les groupements méthyle branchés sur les cytosines sont des marqueurs épigénétiques qui diminuent l’expression des gènes.
- Les divers phénotypes des souris Avy//a peuvent apparaître dans une même portée. Cela suggère que la méthylation de l’ADN d’un gène est un processus aléatoire.
4 - Alimentation maternelle et phénotypes du pelage des souris Avy//a
- Les deux lots de souris de ce document ne diffèrent que par l’alimentation reçue par les mères gestantes de génotype a//a pendant la gestation et la lactation. Les différences entre les phénotypes des deux groupes de descendants Avy//a peuvent donc être attribuées à l’enrichissement de l’alimentation en nutriments favorisant la méthylation de l’ADN reçus par les souriceaux d’un groupe durant leur vie fœtale et durant les 21 jours qui suivent la naissance, avant le sevrage.
- On constate que la fréquence des souris au phénotype tacheté est à peu près la même dans les deux groupes. En revanche, la fréquence des phénotypes témoignant d’une forte expression de l’allèle Avy (jaune + légèrement tacheté) est nettement plus élevée dans le groupe non supplémenté que dans l’autre : 59% contre 33%. Inversement, les phénotypes résultant d’une faible expression de l’allèle Avy (fortement tacheté et pseudo agouti) sont plus fréquents dans le groupe supplémenté.
- L’apport dans l’alimentation maternelle d’un supplément de nutriments contribuant à la méthylation de l’ADN a donc pour effet global d’augmenter la probabilité pour que l’allèle Avy s’exprime peu chez les souris de génotype Avy//a. Connaissant l’effet de la méthylation sur l’intensité d’expression d’un gène, on peut dire que l’alimentation maternelle en augmentant la probabilité de méthylation de la région régulatrice de l’allèle Avy, fait que la probabilité pour que les descendants Avy//a aient un phénotype fortement tacheté ou pseudo agouti est plus grande.
- Cet exemple illustre un nouvel aspect des relations entre génotype, environnement et phénotype, nouveau domaine d’étude en plein essor actuellement. Le fait important est que le marquage épigénétique d’un gène durant la vie fœtale, sous l’effet de facteurs d’environnement, retentit sur le phénotype de l’adulte. La méthylation de l’allèle Avy dans l’exemple du modèle de la souris agouti se réalise très tôt au cours du développement dans les cellules embryonnaires à l’origine des cellules de la peau. Elle se transmet ensuite au cours des mitoses et se retrouve chez les cellules de l’adulte. Il y a donc une hérédité cellulaire des marqueurs épigénétiques. Chez les souris aux phénotypes plus ou moins tachetés, les cellules embryonnaires cutanées ont été différemment méthylées durant la vie embryonnaire et ces différences se retrouvent dans les cellules plus tard, d’où des variations dans l’expression de l’allèle Avy suivant les territoires de la peau.
5 – Des phénotypes cutanés à la susceptibilité à certaines maladies.
- L’allèle Avy s’exprime dans tous les tissus. Un des objectifs de l’expérimentation réalisée par Dolinoy et al. est de voir si l’expression de cet allèle est affectée par la méthylation dans tous les tissus et pas uniquement dans les cellules dermiques sécrétrices de la protéine agouti. Si oui, comment cela se répercute-t-il sur le phénotype ? Au lieu d’utiliser une supplémentation avec des nutriments favorisant la méthylation de l’ADN, ils ont supplémenté les souris gestantes avec un phyto-oestrogène extrait du soja. L’analyse de la fréquence des phénotypes des souris Avy issues du croisement indique que les phénotypes témoignant d’une forte expression de l’allèle Avy sont moins nombreux dans le groupe supplémenté. Autrement dit, l’apport de genistein a eu le même effet que celui de nutriments « méthylinisants ». Le graphique sur la méthylation montre que le genistein a augmenté la probabilité de méthylation de l’allèle Avy et que la méthylation acquise durant la début de la vie persiste dans les tissus adultes, puisqu’elle est la même à 150 jours qu’au 21ème jour. On retrouve la notion fondamentale de la transmission des marques épigénétiques au cours des divisions cellulaires.
- Quand on compare l’évolution du poids des souris de phénotypes extrêmes (jaune et pseudo agouti), on constate que dès la dixième semaine, le poids moyen des souris jaunes est nettement supérieur à celui des souris agouti : 35g contre 25g ; cette différence s’accroît par la suite : 55g contre 35 à la soixantième semaine. Dans tous les tissus, l’allèle Avy est très peu méthylé chez les souris jaunes et fortement méthylé chez les souris pseudo agouti. La différence de méthylation de l’allèle Avy se répercute non seulement sur la couleur du pelage mais aussi sur le caractère poids de l’animal. La méthylation de l’allèle Avy réduit son expression au niveau des régions du système nerveux impliqué dans la prise alimentaire et dans le tissu adipeux. Il en résulte un poids plus faible que chez les souris jaunes où l’allèle Avy peu ou pas méthylé a une forte expression dans ces tissus, ce qui entraîne une obésité.
- On constate toutefois qu’il n’y a pas de différence significative dans l’évolution du poids des phénotypes allant du jaune au fortement tacheté. Seule une très forte méthylation de l’ADN en inhibant l’expression de l’allèle Avy a des répercussions sur le poids de la souris.
- Puisque le genistein augmente la probabilité de méthylation de l’allèle Avy et donc la fréquence du phénotype pseudo agouti, on voit que cette substance a une influence non seulement sur la couleur du pelage mais aussi sur le poids : les souris pseudo agouti ne sont pas obèses. Cet exemple supporte l’idée que l’alimentation maternelle pendant la gestation ou peu après la naissance peut avoir des répercussions sur des aspects phénotypiques de l’adulte, par exemple favoriser l’apparition d’une obésité qui peut contribuer au déclenchement d’une maladie métabolique comme le diabète. Mais bien entendu, il faut être prudent quand on veut transférer à l’espèce humaine les conclusions du modèle de la souris agouti. En particulier, on n’a pas mis en évidence dans le génome humain la présence d’un allèle d’un gène de susceptibilité à l’obésité susceptible d’une méthylation variable suivant les individus et influencée par l’environnement.
- Le document sur le bisphénol montre que cette substance augmente la fréquence des phénotypes où l’allèle Avy est peu ou pas méthylé. Il a donc un effet opposé au genistein. On peut alors penser qu’il augmente la probabilité de souris obèses à l’état adulte.