
Famille multigénique des opsines
Informations scientifiques (Rappel)
La rétine humaine est un tissu nerveux qui renferme notamment des cellules photoréceptrices de deux types : les cônes et les bâtonnets. Les cônes sont responsables de la vision des couleurs ; les bâtonnets ne permettent pas la vision des couleurs mais sont très sensibles à la lumière et permettent une vision en très faible éclairement.
La vision des couleurs est liée à l’existence dans la rétine de trois types cellulaires (les cônes) qui sont des photorécepteurs synthétisant trois pigments, des opsines, qui présentent des niveaux d’absorption différents dans le rouge (opsine rouge ou L), le vert (opsine verte ou M) ou le bleu (opsine bleue ou S).
Avec un seul des pigments, on ne voit pas les couleurs ; la présence des trois pigments permet une vision trichromatique.
Les pigments des cellules photoréceptrices
Toutes ces cellules contiennent un pigment de nature essentiellement protéique, excitable par des photons, qui varie selon le type de cellules photoréceptrices :
-
les bâtonnets synthétisent un pigment appelé rhodopsine (constitué d’une opsine et d’un groupement prosthétique, le 11-cis-rétinal lié à la lysine 216 de la protéine, dont le précurseur est le all-trans-rétinol ou vitamine A. Un déficit alimentaire en vitamine A conduit ainsi à l’héméralopie ou cécité nocturne). La rhodopsine présente une bande d’absorption large dans la région visible du spectre avec un pic à 500 nm.
-
Les cônes synthétisent un pigment appelé opsine ; il existe trois types de cônes en fonction du pigment qu’ils synthétisent : les cônes à opsine S (Short ou bleue), opsine M (Medium ou verte) et opsine L (Long ou rouge), ces différentes opsines différant par leur spectre d’absorption.
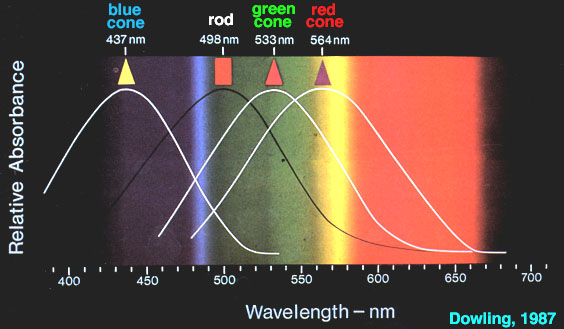
Les spectres d’absorption des trois opsines photoréceptrices ont été mesurés en éclairant des cônes avec un faisceau de lumière de 1 mm seulement. Les réponses des différents cônes montrent trois groupes : certaines sont excitées au maximum par la lumière bleue, d’autres sont excitées au maximum par la lumière verte et les dernières par la lumière orangée-rouge.
Les cônes S contiennent une opsine ayant un maximum d’absorption dans les courtes longueurs d’onde du visible (« opsine S ou bleue »), les cônes M contiennent une opsine ayant un maximum d’absorption dans les longueurs d’onde moyennes du visible (« opsine M ou verte »), les cônes L contiennent une opsine ayant un maximum d’absorption dans les grandes longueurs d’onde du visible (« opsine L ou rouge »).
Bien entendu, tous les photorécepteurs possèdent les mêmes gènes codant pour les opsines, mais un seul de ces gènes s’exprime dans un photorécepteur donné.
Les protéines photoréceptrices des cellules en cône sont, comme l’opsine de la rhodopsine, des récepteurs à sept hélices, et elles contiennent elles aussi le 11-cis-rétinal.
Localisation chromosomique des gènes codant pour les opsines humaines
Les gènes codant pour les opsines M ou verte et L ou rouge sont situés sur le chromosome X. Le gène codant pour l’opsine S ou bleue est porté par le chromosome 7.
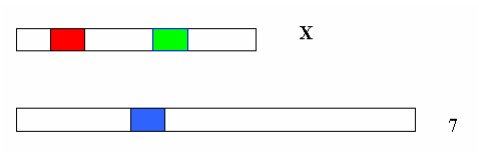
Chez les Primates, la vision trichromatique n’est possible que dans certains groupes :
-
les Singes de l’Ancien Monde (Afrique, Asie et Europe), ou catarhiniens, possèdent, comme l’Homme, le gène de l’opsine S (bleue) porté par un autosome ; le gène de l’opsine M (verte) porté par le chromosome X ; le gène de l’opsine L (rouge) porté par le chromosome X. Ces Singes sont donc tous trichromates ;
-
les Singes du Nouveau Monde (Amérique), ou platyrhiniens, possèdent : le gène de l’opsine S (bleue), situé sur un autosome et un seul autre gène codant pour une opsine S/M, porté par le chromosome X.
Pistes d’exploitation pédagogique
L’exploitation des données fournies permet d’aborder la notion d’apparition de nouveaux gènes par duplication génique suivie de mutations différentes. Les nouveaux gènes codent ici pour des protéines qui, tout en ayant conservé la même fonction globale (absorption de radiations lumineuses), présentent des propriétés légèrement différentes (les longueurs d’ondes absorbées par les différents pigments ne sont pas les mêmes).
Plusieurs arguments suggèrent l’idée d’une origine commune pour les gènes des opsines :
-
les opsines présentent de nombreuses similitudes : leurs propriétés et leurs structures se ressemblent. Elles sont toutes impliquées dans la conversion de la lumière en mouvement ionique puis en signal nerveux. Chaque molécule photosensible contient sept hélices alpha transmembranaires ;
-
les séquences d’acides aminés des opsines absorbant le vert et des opsines absorbant le rouge sont particulièrement semblables. Trois résidus localisés près du rétinal déterminent la différence spectrale entre les récepteurs pour le vert et pour le rouge. Le remplacement d’un résidu polaire par un résidu non polaire (par exemple, la sérine par l’alanine) à chacune de ces positions déplace lmax (Intensité lumineuse) vers le rouge d’environ 10 nm. Trois acides aminés expliquent la majeure partie de la différence de 30 nm entre les maxima d’absorption ;
-
les séquences nucléotidiques ou les séquences peptidiques des opsines humaines présentent des similitudes importantes.
Fichiers des séquences
-
Opsines - ADN.edi : séquences nucléiques strictement codantes des gènes des opsines humaines rouge, verte et bleue ;
-
Opsines - Protéines.edi : séquences protéiques des opsines rouge, verte et bleue humaines.
Comparaison en 3D
Représentation en rubans des trois ospines humaines (document obtenu avec le logiciel RasTop).
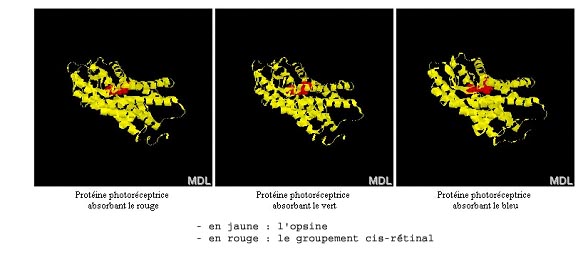
Les similitudes constatées sont un argument en faveur d’une origine commune de ces protéines, donc des gènes qui les codent.
La comparaison de séquences protéiques et/ou nucléiques permet de dégager la notion de molécules homologues (protéines homologues ou gènes homologues) grâce à l’importance des similitudes constatées. Le relevé des différences permet de préciser le degré de parenté entre les protéines et/ou les gènes, et donc d’établir une phylogénie pour ces gènes.
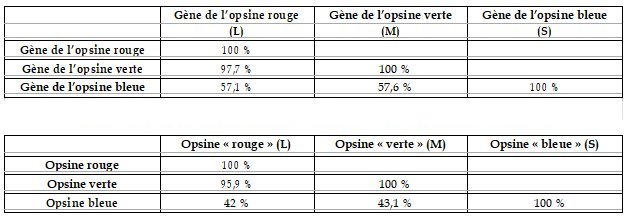
Matrice des identités créée à partir d’un alignement avec discontinuité des opsines (séquences nucléiques en haut et protéiques en bas)

Arbre phylogénétique obtenu à partir de l’exploitation des matrices des identités.
La phylogénie obtenue permet de discuter des mécanismes à l’origine de l’apparition de gènes différents à partir d’un seul.
Datation des duplications
La prise en compte d’observations réalisées chez d’autres Primates permet de dater approximativement la dernière duplication génique : seuls les Singes de l’Ancien Monde possèdent trois gènes d’opsine et notamment un gène d’opsine rouge et un gène d’opsine verte. Les Singes du Nouveau Monde ne possédant que deux gènes, on peut dire que la duplication à l’origine des gènes des opsines verte et rouge à partir d’un même gène ancestral a dû avoir lieu dans la lignée menant aux Singes de l’Ancien Monde, et a donc dû se produire après la séparation de cette lignée de celle des Singes du Nouveau Monde, soit il y a environ 40 à 23 Ma.
Translocation de gènes
Les gènes des opsines provenant tous d’un même gène ancestral, mais étant situés sur deux chromosomes différents, il faut imaginer un phénomène de translocation d’un des deux exemplaires de gènes lors de la duplication du gène ancestral.
L’histoire évolutive de la famille des opsines
On peut demander aux élèves de compléter l’arbre phylogénétique à l’aide des informations fournies par la comparaison des gènes des opsines comme ci-dessous :
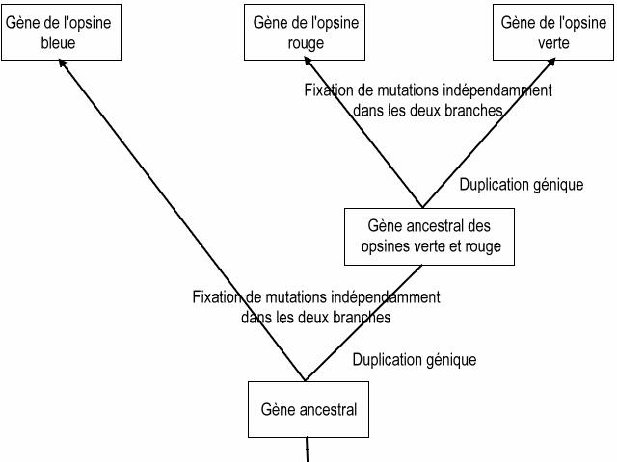
Reconstitution de l’histoire de la famille des opsines.