
Les médiateurs de l’inflammation
La présence de signaux de danger (DAMP) ou de motifs pathogéniques (PAMP) est perçue par les cellules immunitaires via leurs PRR.
Les cellules présentes dans le tissu infecté ou lésé, telles que les phagocytes mononucléés résidents (macrophages et cellules dendritiques) et les mastocytes, sont les premières cellules activées par des signaux de dangers. En réponse à cette activation, elles libèrent de l’histamine, des cytokines pro-inflammatoires et d'autres composés actifs que nous regrouperons ici sous le terme général de médiateurs de l’inflammation. Les conséquences fonctionnelles de cette activation sont l’élimination du pathogène (par ex. par phagocytose) et/ou la réparation de la lésion (remodelage de la matrice extracellulaire).
|
A. Homéostasie 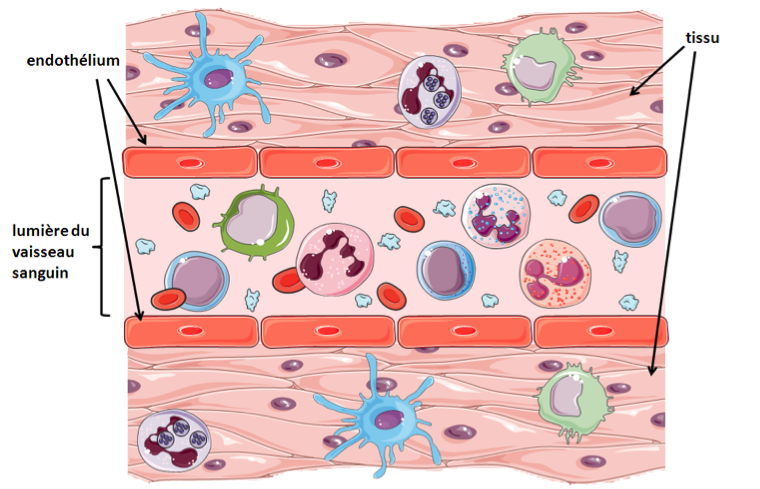 |
B. Inflammation
|
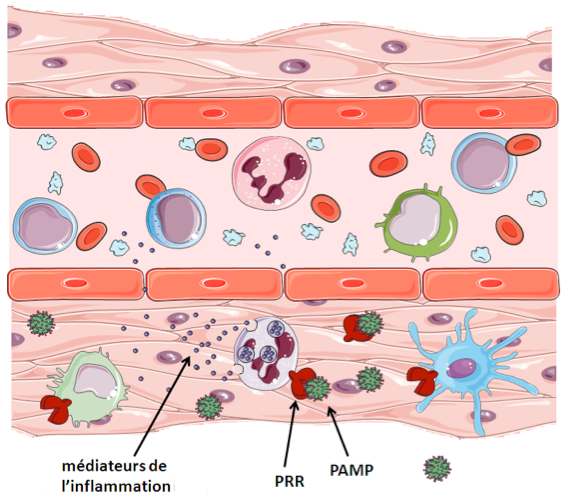
Mise en place d’une réponse inflammatoire
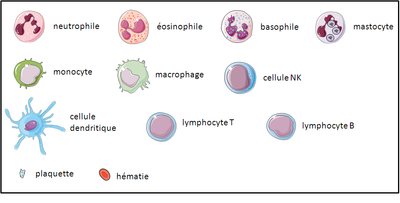
A - Les cellules immunitaires résidantes des tissus sont les mastocytes, les macrophages et les cellules dendritiques. Les cellules de l’immunité qui patrouillent dans l’organisme via le sang sont les neutrophiles, les éosinophiles, les basophiles, les monocytes, les cellules NK et les lymphocytes T et B.
B - Lors d’une infection, la présence de pathogène est détectée par ces cellules via leurs PRR, récepteurs reconnaissants les PAMP, qui sont des motifs particuliers aux agents infectieux. Les cellules immunitaires ainsi activées libèrent des médiateurs de l’inflammation dans le milieu extracellulaire, dont les effets combinés vont permettre le recrutement de cellules circulantes, l’élimination du pathogène et la réparation de la lésion.
L’histamine
L’histamine est une amine vasoactive stockée dans les granules des mastocytes et des basophiles. Les mastocytes activés par la reconnaissance de DAMP ou de PAMP ou par les anaphylatoxines C3a et C5a, libèrent l’histamine stockée dans leurs granules dans le milieu extracellulaire.
L’histamine augmente la perméabilité vasculaire et permet la contraction des muscles lisses localement. Elle agit aussi au niveau de système nerveux et provoque la sensation de douleur.
Les cytokines pro-inflammatoires
Les cytokines (du grec « cyto » : cellules et « kinos » : mouvement) sont des petites protéines sécrétées par les cellules en réponse à divers stimuli. Au niveau de la réponse immunitaire, elles permettent la communication entre les cellules immunes et l’orientation de la réponse en fonction de la nature du signal détecté.
- TNFα (Tumor Necrosis Factor α)
Cette cytokine est produite par les macrophages, les cellules dendritiques résidentes et les mastocytes. Ces derniers renferment dans leurs granules des stocks préformés de TNFa.
Le TNFa stimule l’expression de molécules d’adhérence et la production de chimiokines par les cellules endothéliales permettant le recrutement des leucocytes sanguins (neutrophiles, éosinophiles, monocytes ou les NK) vers le foyer inflammatoire.
Le TNFa active aussi les systèmes microbicides des phagocytes et est mitogène pour les lymphocytes T et B (pour la mise en place de la réponse adaptative si la réponse innée n’est pas suffisante à la résolution de l’inflammation).
Enfin, le TNFα active la production de facteurs de croissance, qui seront indispensables à la réparation du tissu endommagé.
- IL-1 (Interleukine-1)
L’IL-1 est sécrétée par de nombreuses cellules après la reconnaissance de PAMP : les leucocytes, mais aussi les cellules endothéliales, les fibroblastes, présents dans tous les tissus. L’IL-1 provoque des effets locaux et des symptômes plus généraux (effets systémiques) permettant à l’organisme de combattre la maladie.
Comme le TNFα, l’IL-1 stimule l’expression de molécules d’adhésion au niveau de l’endothélium vasculaire favorisant ainsi la migration des leucocytes circulants vers le site enflammé.
Un de ses rôles le plus important est l’induction de la fièvre. L’IL-1 agit directement sur les centres de l’hypothalamus qui contrôlent la température corporelle. L’augmentation de la température du corps donne un avantage à l’organisme sur le pathogène : les lymphocytes se divisent plus vite et le système immunitaire adaptatif est donc mobilisé plus rapidement alors que certains pathogènes ne se divisent pas ou moins bien à des températures élevées. L’IL-1 induit aussi une diminution de l’appétit et favorise un certain type de sommeil.
- IL-6 (Interleukine-6)
L’interleukine-6 est produite par les phagocytes (macrophages et cellules dendritiques) et les cellules endothéliales en cas d’inflammation. Elle induit localement l’activation des phagocytes et la modification de l’endothélium. Elle favorise le recrutement de monocytes sanguins vers les tissus et la production de protéines de la phase aiguë par les hépatocytes.
- Autres cytokines
Le TNFα, l’IL-1 et l’IL-6 sont les principales cytokines de l’inflammation.
L’activation des PRR stimule aussi la production d’autres cytokines, dont les IFN (interférons) qui induisent la production de molécules antivirales, ainsi que l’IL-12 et l’IL-18, cytokines qui vont activer les cellules NK (Natural Killer), cellules de l’immunité innée. Ces mécanismes jouent un rôle capital en freinant la diffusion des infections virales.
Les chimiokines
Les chimiokines sont des cytokines impliquées dans la migration cellulaire. Elles sont sécrétées par de nombreuses cellules sentinelles après stimulation par des signaux de dangers. Elles forment alors un gradient, et dirigent ainsi la migration des populations leucocytaires requises vers le tissu enflammé (figure ci-dessous, vidéos "Recrutement des leucocytes" et "Chimiotactisme neutrophile").
On répartit les chimiokines en deux classes, en fonction de la distance séparant deux résidus cystéines conservés : les chimiokines α qui ont une séquence CXC (cystéine-résidu aléatoire-cystéine) et les chimiokines β qui ont une séquence CC (cystéine-cystéine).
La plupart des chimiokines intéragissent avec plusieurs récepteurs, et chaque récepteur peut reconnaitre plusieurs chimiokines. Chaque population leucocytaire exprime un groupe défini de récepteurs, ce qui permet aux différents types de cellules immunitaires de répondre spécifiquement à certaines chimiokines seulement.
|
Chimiokines
|
Cellules sources
|
Récepteurs
|
Cellules cibles
|
|
CXCL8
|
monocytes, macrophages, endothélium, fibroblastes, kératinocytes |
CXCR1 et CXCR2
|
neutrophiles
|
|
CXCL10
|
monocytes, endothélium, fibroblastes, kératinocytes, lymphocytes T |
CXCR3
|
monocytes, cellules NK
|
|
CXCL1, 2 et 3 |
monocytes, endothélium, fibroblastes
|
CXCR2
|
neutrophiles
|
|
CCL2
|
monocytes, macrophages, fibroblastes, kératinocytes
|
CCR1 et 2
|
macrophages, basophiles
|
|
CCL5
|
lymphocytes T, endothélium, plaquettes
|
CCR1, 3 et 4
|
basophiles, lymphocytes T
|
Par exemple, les neutrophiles expriment CXCR1 et 2, ce qui leur permet de répondre aux chimiokines CXCL8, 1 et 2.
Autres médiateurs de l’inflammation
- Les médiateurs lipidiques de l’inflammation
Lorsque qu’une cellule (phagocyte ou mastocyte) détecte un signal de danger, la phospholipase A2 est activée (via la stimulation par le TNFα). Cette enzyme transforme les acides gras de la membrane plasmique de la cellule en leucotriènes, prostaglandines et Platelet Activating Factor (PAF). Ces composés sont chimiotactiques pour les neutrophiles et les macrophages, et induisent aussi l’augmentation de la dilatation des vaisseaux et leur perméabilité, facilitant l’arrivée des leucocytes sur le site de l’inflammation.
- Les protéines de la phase aigüe
Elles sont produites par les hépatocytes (cellules du foie) au moment de la phase aigüe de l’inflammation. Nous donnerons ici deux exemples, les deux pentraxines CRP (C-Reactive Protein) et SAP (Serum Amyloid-P). Toutes les deux sont de puissantes opsonines favorisant la phagocytose des pathogènes par les macrophages.
La CRP est produite rapidement en réponse à l’IL-6, et son dosage dans le serum est utilisé en clinique pour diagnostiquer une inflammation.
- Le système des kinines
La bradykinine est produite à la suite de l’activation du facteur de Hageman (ou facteur XII) du système de coagulation sanguine, lorsqu’il rencontre du collagène après une lésion vasculaire (le collagène est exposé au contact du sang après désorganisation du tissu endothélial). Ce peptide est un vasoactif très puissant qui induit la dilatation des veinules, l’augmentation de la perméabilité vasculaire et la contraction locale des muscles lisses, évènements favorisants le recrutement de leucocytes.
Comme l’histamine, les prostaglandines et la bradykinine participent au déclenchement de la sensation douloureuse.
- Les enzymes et le système de la plasmine : remodelage tissulaire
Le lysozyme produit par les macrophages dégrade les protéoglycanes de la paroi bactérienne (directement accessible sur les bactéries Gram+). L’urokinase, la collagénase, l’élastase, ainsi que les métalloprotéinases (MMP) détruisent la matrice extracellulaire, ce qui favorise le remodelage tissulaire, et facilitent ainsi la migration des cellules immunitaires vers le site inflammé.
L’urokinase permet le clivage du plasminogène (protéine présente dans le plasma mais aussi les tissus) en plasmine. Cette dernière induit le remodelage tissulaire (dissout la fibrine, remodèle le collagène), et favorise l’angiogénèse en déclenchant la libération de cytokines qui induisent la prolifération et la migration des cellules endothéliales.
- Les protéines microbicides et les dérivés oxygénés
Les défensines et cathélicidines sont de petits peptides chargés positivement, capable de s’insérer dans les membranes contenant des phospholipides de charge négative (comme les bactéries Gram-), et de former ainsi des canaux perméables capables de lyser les pathogènes (bactéries, champignons, et même virus comme Herpes simplex). La lactoferrine est libérée par les neutrophiles activés. Elle séquestre le fer et en prive donc les pathogènes, ce qui inhibe leur croissance.
Les cellules phagocytaires activées produisent des dérivés toxiques de l’oxygène (eau oxygénée, oxygène singulet) et détruisent ainsi le pathogène phagocyté.
Elles sécrètent aussi de l’oxyde nitrique (NO) toxique pour les bactéries extracellulaires.
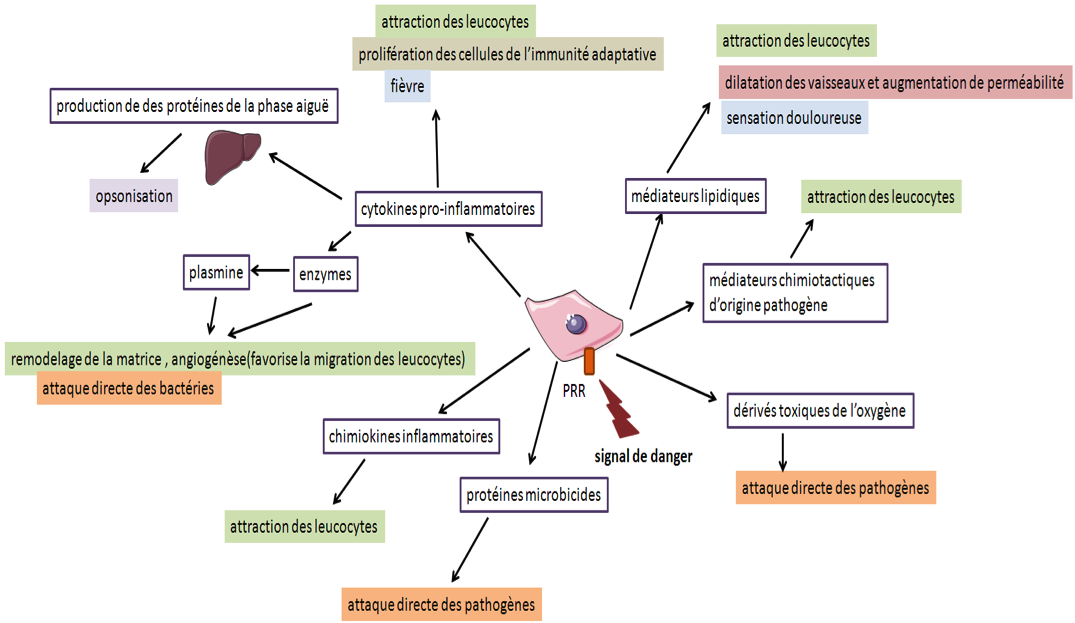
Les médiateurs de l’inflammation