
Structures impliquées dans la genèse du sommeil paradoxal
Le sommeil paradoxal correspond au dernier stade d'un cycle de sommeil (=stade v). Il n'apparait qu'en sommeil lent. Il se caractérise par:
-
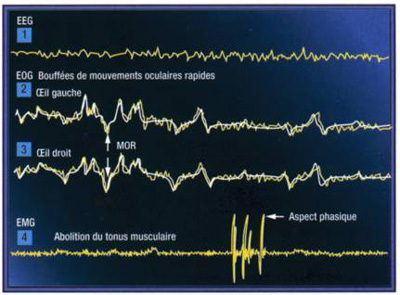
une activité corticale caractérisée par un EEG présentant une activité de bas voltage et de haute fréquence qui rappelle l'EEG de veille (=désynchronisation corticale) mais qui est différent de celui-ci
-
des mouvements occulaires (=REM= Rapid eyes movements)
NB: en anglais le sommeil paradoxal est appelé REM-sleep et le sommeil lent non-REM-sleep
-
des « Pointes ponto-genouillées occipitales » (=PGO): la désynchronisation corticale observable pendant le SP s’accompagne d’ondes lentes sporadiques que l’on enregistre à partir d’électrodes placées au niveau du pont, des corps genouillés latéraux et du cortex occipital
-
une atonie musculaire
On n'a pas d'idée précise, à l'heure actuelle, sur la relation existant entre les centres du sommeil lent (et de l'éveil) et ceux impliqués dans le sommeil paradoxal. Pourtant il y a forcément une relation puisque le sommeil paradoxal n'apparaît qu'en sommeil lent. Cet article se limitera à une description des principaux réseaux de neurones impliqués dans le sommeil paradoxal.
La mise en évidence de l'importance du tronc cérébral a été faite à partir d'expérience de sections réalisées chez le chat
-
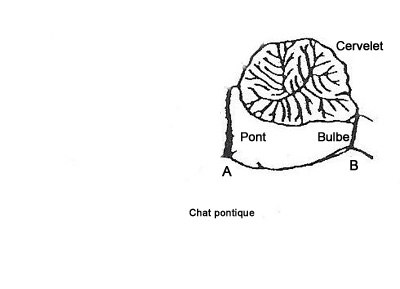
Etude de chat pontique
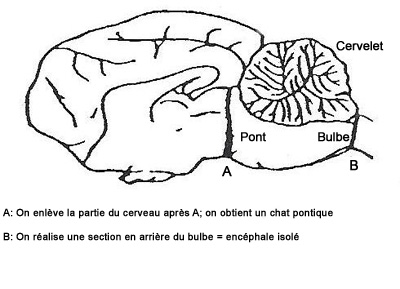
Figure 2. Localisation des sections réalisées pour obtenir un "chat pontique" (section en A) ou un encéphale isolé (section en B)
Lorsqu' on enlève la partie du cerveau située en avant du pont chez un chat, il continue à présenter périodiquement les symptômes du sommeil paradoxal. Un tel animal peut survivre plusieurs mois à condition que son équilibre thermique, nutritionnel et hydrominéral soit assuré artificiellement . L'animal subit alors, avec une régularité d'horloge, des périodes d'atonie posturale. Ce phénomène peut paraître étonnant puisque les animaux décérébrés manifestent en général une hypertonie importante appelée hypertonie de décérébration. Ces périodes d'atonies s'accompagnent de mouvements oculaires rapides en relation avec une activité pontogenouillée occipitale (PGO) au niveau du pont. Elles ont, de plus, une durée similaire à celles des périodes de SP chez l'animal intact.
Le sommeil paradoxal prend donc naisssance dans le pont.
-
Chat avec encéphale isolé
Une section de la moelle cervicale, juste en arrière du bulbe (= encéphale isolé, section en B) ne supprime pas les signes électiques du sommeil paradoxal en avant de la section. L'atonie musculaire qui dépend des motoneurones est évidemment supprimée par la lésion
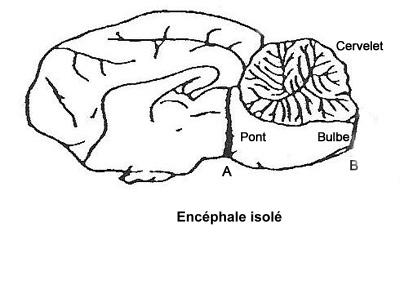
Ces deux expériences permettent de conclure que le rhombencéphale (= bulbe rachidien +cervelet + pont) est suffisant pour mettre en jeu le someil paradoxal.
D'autres expériences montrent que la stimulation par le pont est à elle seule insuffisante pour déclencher le sommeil paradoxal. Il faut donc supposer des interactions réciproques complexes entre le pont et le bulbe pour déclencher le sommeil paradoxal. De plus le contrôle du sommeil paradoxal semble assuré par le cerveau antérieur.
Nous allons maitnenant préciser quels sont les réseaux neuronaux qui sont mis en jeu lors du sommeil paradoxal et comment ils permettent l'atonie musculaire et l'activation corticale.
Il est formé par des neurones dit SP-OFF car ils sont inactifs pendant le Sommeil Paradoxal. Pendant l'éveil ils empêchent les neurones du réseau exécutif du sommeil paradoxal (voir plus loin) de fonctionner. C'est leur inhibition qui permet l'installation du sommeil paradoxal.
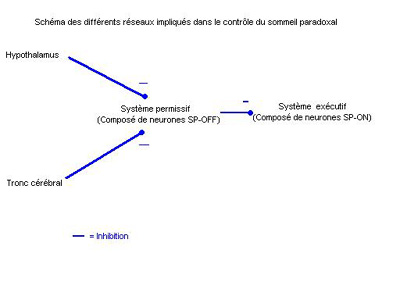
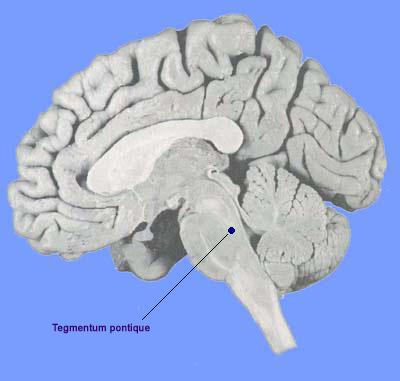
Ils sont localisés entre autre dans le tegmentum pontique. Ce sont des neurones aminergiques (à noradrénaline, à sérotonine ou à histamine). Ces neurones sont inhibés par des neurones de l'hypothalamus et du tronc. Certaines hypersomnies résulteraient de l'arrêt prolongé du système permissif.
Il est formé par des neurones dit SP-ON car ils sont actif pendant le sommeil paradoxal (mais pas pendant le sommeil lent). Les différentes composantes du SP détaillées en introduction dépendent chacune de groupes de neurones SP-ON spécifiques.
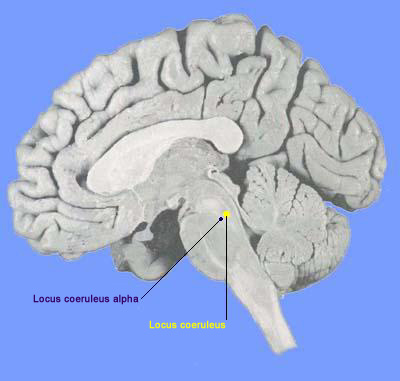
La destruction du locus coeruleus alpha empêche l'atonie musculaire observée pendant le sommeil paradoxal. Chez ces animaux, la désynchronisation corticale, les mouvements rapides des yeux et les PGO après un stade 4 de sommeil à ondes lentes, attestent d'une phase de sommeil paradoxal. Cependant les animaux ont simultanément un comportement moteur à la place de l'atonie normale. Les comportements observés sont caractéristiques de l'espèce: attaque prédatrice, rage (dos rond, érection des poils), peur, toilettage, exploration. Les mouvements sont très fortement liés aux pointes PGO. Tous ces comportements ne sont jamais dirigés vers un élément de l'environnement et l'animal est largement insensible à tout stimulus externe. Les animaux "vivent" leur rêve.
Il convient tout d'abord de souligner que l'EEG enregistré pendant le sommeil paradoxal, même s'il fait penser à celui de l'éveil, est différent de celui-ci.
Le tegmentum pontique contient des neurones SP-OFF (inactifs pendant le SP) qui permettent au sommeil paradoxal de s'exprimer (pendant l'éveil ils empêchent les neurones SP-ON de s'exprimer) lorsqu'ils sont inhibés. Il contient également des neurones SP-ON (actifs pendant le SP mais pas pendant l'éveil ni le SL). Ces derniers sont cholinergiques mais contiennent sans doute d'autres neurotransmetteurs. Ces neurones agissent sur le cortex et entraînent la désynchronisation transitoire des neurones du cortex. Ils ont également une action inhibitrice sur les noyaux réticulaires du thalamus. Ils permettent l'installation du sommeil paradoxal.
Les neurones cholinergiques du tegmentum pontique dorsal (neurones SP-ON) sont donc responsables à la fois de l'initiation et du maintien du SP. Néanmoins différentes manipulations montrent qu'ils ne sont pas suffisants pour l'apparition du SP:
-
Si on augmente la transmission de l'Acétylcholine on augmente l'éveil mais aussi le sommeil paradoxal au détriment du sommeil lent.
-
L'introduction de Carbachol (agoniste cholinergique) dans la formation réticulée pontique entraîne une augmentation de 300 % du SP
On peut schématiser les interactions entre les différents réseaux étudiés par l'animation suivante (power point).