
Le traitement cérébral de l'information visuelle
Les aires visuelles
-
l'aire V1 ou cortex visuel primaire ou aire striée (aire 17 de Brodmann)
-
les aires extrastriées qui correspondent à l'ensemble des aires visuelles autres que V1 (c'est à dire V2, V3, VP= V3 centrale, V4, MT......)
Dans les aires extrastriées, les aires préstriées correspondent aux aires visuelles associatives, ce sont toutes les aires impliquées dans le traitement des informations visuelles, autres que V1 et V2.
Dès les années 1970, l'étude du singe- hibou et du macaque a montré que le cortex visuel d'association (cortex préstrié) est constitué de plusieurs aires cérébrales séparées de l'aire V1 par l'aire V2. A l'aide d'électrodes, on a pu enregistrer l'activité électrique de cellules du cortex préstrié de singes à qui on présentait différents stimuli : couleurs, barre d'orientation variable, points lumineux en mouvement ou se déplaçant dans différentes directions.
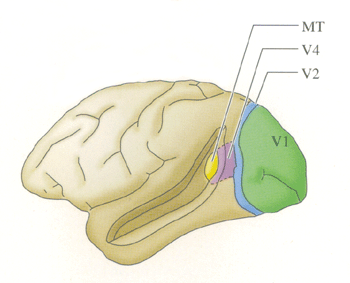
Ainsi a été découvert l'aire MT ou aire V5 chez l'homme. C'est une région dont toutes les cellules sont sensibles au mouvement et pour la plupart , spécifiques de certaines directions, et dont aucune n'est sensible à la couleur des cibles en mouvement.
Une autre région, l'aire V4, a été identifiée. Ses cellules sont spécifiques de certaines longueurs d'onde lumineuses et la plupart d'entre elles sont également sensibles à l'orientation.
La plupart des cellules de deux autres aires voisines, les aires V3 et V3A (chez l'homme) sont sensibles aux formes, mais, comme les cellules de l'aire V5, elles ne sont pas sensibles à la couleur des stimuli.
Chez l'homme, c'est grâce à des études en imagerie cérébrale fonctionnelle (notamment par une approche nommée IRMF rétinotopique ou IRMf Fourier), qu'une disposition comparable des aires visuelles a été montrée. A l'aide de stimuli ponctuels, présentés en des endroits définis de la rétine, on est parvenu à localiser au moins dix représentations distinctes du champ visuel. Les études cliniques de déficits visuels successifs à des lésions du cortex extrastrié viennent confirmer cette spécialisation fonctionnelle du cortex extrastrié humain.
Toutes ces aires visuelles bilatérales, présentent en effet une organisation rétinotopique; elles montrent une correspondance de la position de leurs cellules avec celles des différentes zones (champs récepteurs) d'un hémichamp (gauche si elles appartiennent à l'hémisphère droit et réciproquement), cependant les champs récepteurs des cellules des aires préstriées, s'ils sont très spécifiques de certaines caractéristiques de l'espace visuel, sont beaucoup plus larges que ceux des cellules des aires V1 et V2.
 |
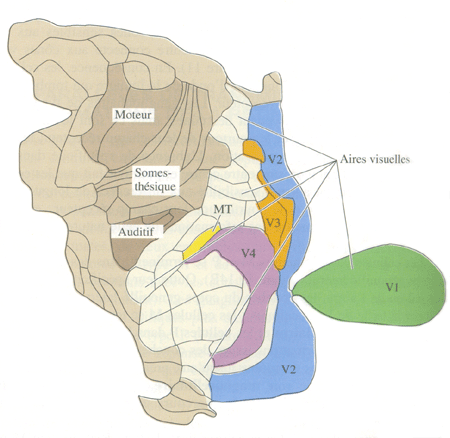 |
| Figure 1. Aires visuelles du singe (Neurosciences Purves) | Figure 2. Carte des aires visuelles du singe (Neurosciences, Purves) |
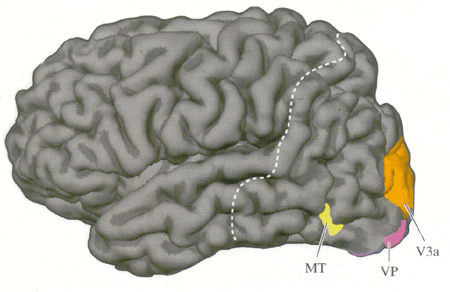
Ainsi lorsqu'une personne regarde une scène abstraite colorée, la région du cerveau où le débit sanguin augmente le plus est le gyrus fusiforme (qui correspond à l'aire V4 du macaque). De même lorsqu'une personne regarde une image de mouvement en noir et blanc, la zone de plus forte augmentation de débit sanguin est plus latérale et bien séparée de V4, c'est l'aire V5 (aire MT du singe). Cependant, dans les deux cas le débit sanguin augmente aussi dans l'aire V1 et l'aire V2.
 |
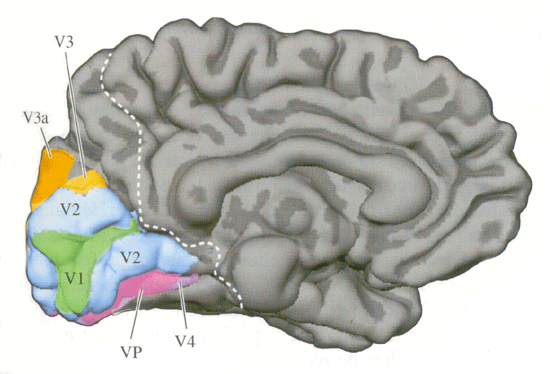 |
| Figure 3. Aires visuelles humaines, face externe (Neurosciences, Purves) | Figure 4. Aires visuelles humaines, face interne (Neurosciences, Purves) |
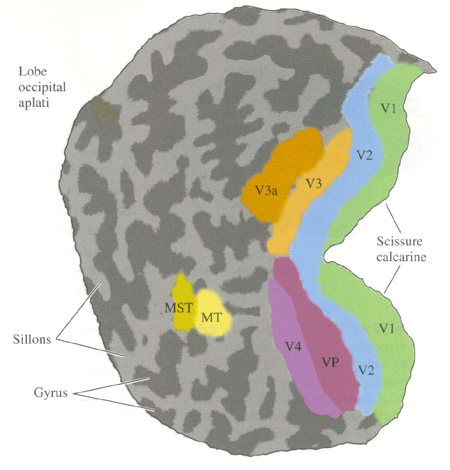 |
| Figure 5. Cartes des aires visuelles humaines (Neurosciences, Purves) |
Des aires V1 et V2 aux aires préstriées: aires visuelles associatives
Les capacités de distribution de l'information de ces aires vers les aires préstriées résultent de leur organisation anatomique et fonctionnelle.
L'aire V1 (où les cellules les plus actives peuvent être révélées par un colorant spécifique de la cytochrome-oxydase mitochondriale) est métaboliquement divisée en colonnes de cellules s'étendant de la surface du cortex jusqu'à la substance blanche sous-jacente. Sur des coupes parallèles à la surface, ces colonnes (cylindres fushia sur le schéma ci-dessous) apparaissent comme des taches (blobs) séparées par des régions moins colorées. Margaret Livingstone et David Hubel ont montré que les cellules de l'aire V1 sensibles à la couleur sont concentrées dans ces tâches, tandis que les cellules sensibles à la forme sont entre ces tâches. Ces colonnes sont particulièrement nettes dans les couches 2 et 3 de l'aire V1, qui reçoivent les informations provenant des couches parvocellulaires du corps genouillé latéral. Ces cellules sont sensibles de façon tonique aux stimulis visuels et la majorité d'entre elles code la couleur.
Des structures différentes apparaissent dans la couche 4B de l'aire V1 qui reçoit les informations des couches magnocellulaires du corps genouillé latéral. Ces cellules réagissent de façon transitoire (phasique) aux stimuli visuels, elles sont insensibles à la couleur. La couche 4B de l'aire V1 se projette vers les aires V3 (forme) et V5 (mouvement). Les cellules de la couche 4B qui se connectent à l'aire V5 sont elles-mêmes regroupées dans de petites zones spécifiques!
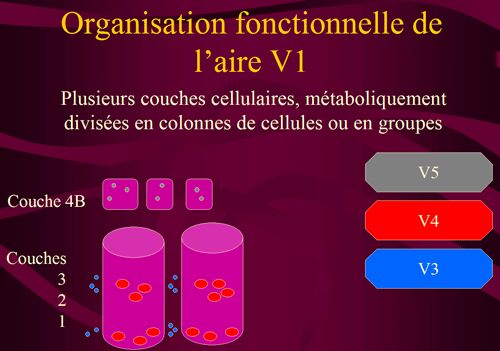 |
| Figure 6. Organisation cellulaire de l'aire V1 (représentation avec la surface corticale vers le bas) Cellules sensibles à la couleur en rouge (colonnes ou "taches" couches 2 et 3), cellules sensibles à la forme en bleu (inter-taches couche 2 et 3) et cellules sensibles au mouvement en gris (couche 4B). |
L'aire V2 est composée de bandes minces et de bandes épaisses, séparées par des régions claires. Les cellules sensibles à la couleur sont regroupées dans les bandes minces et les cellules sensibles à la direction du mouvement sont dans les bandes larges. Des cellules sensibles à la forme se trouvent à la fois dans les bandes larges et dans les espaces clairs entre les bandes.
 |
|
Figure 7. Organisation cellulaire de l'aire V2 |
Les cellules des aires visuelles V1 et V2 ont de petits champs récepteurs et n'enregistrent que certaines caractéristiques des stimuli apparaissant dans leurs champs. Les aires V1 et V2 découpent le champ visuel et envoient des signaux spécialisées en terme de couleur, forme ou mouvement (mais conservant leur topographie) aux aires d'association.
La forme, la couleur et le mouvement
Dans l'état actuel des connaissances, on peut dire qu'il existe 4 systèmes qui traitent en parallèle les diverses caractéristiques d'un objet. L'un détecte le mouvement, un autre la couleur, et deux la forme.
-
la couleur est perçue lorsque les cellules sensibles, présentes dans les colonnes de l'aire V1, envoient des signaux vers l'aire spécialisée V4 et vers les bandes minces de l'aire V2 qui sont connectées à cette dernière
-
la détection des formes colorées résulte d'échanges de signaux entre les régions inter-taches de V1, les régions inter-bandes de V2 et l'aire V4
-
la détection du mouvement et des formes en mouvement se fait lorsque les aires V3 et V5 reçoivent des signaux en provenance directe de la couche 4B de l'aire V1 ou par l'intermédiaire des bandes larges de V2
Voir : le système de la couleur depuis la rétine jusqu'à l'aire V4
Voir : les deux systèmes de la forme depuis la rétine jusqu'aux aires V3 et V4
Voir : le système du mouvement depuis la rétine jusqu'aux aires V3 et V5
On peut noter que la perception de la couleur par un humain se fait 80 millisecondes avant celle du mouvement (le centre de la couleur termine son travail bien avant le centre du mouvement). Sachant que les signaux les plus rapides atteignent le cortex en 30 millisecondes, une telle différence est très importante.
L'unité du monde visuel
Comment les aires spécialisées interagissent-elles pour fournir finalement une image unifiée, préalable indispensable à une prise de conscience de l'environnement visuel?
L'hypothèse la plus simple serait que les aires spécialisées communiquent finalement le résultat de leurs opérations à une aire unique qui synthétiserait toutes ces informations. Mais il n'a jamais été trouvé de structure anatomique à laquelle toutes les aires spécialisées seraient connectées. Les aires spécialisées se connectent plutôt entre elles.
Ainsi les aires V4 et V5 se connectent entre elles directement et réciproquement. Par ailleurs elles projettent aussi toutes les deux vers les régions pariétales et temporales du cerveau, mais on a montré récemment que ces projections aboutissent dans des zones différentes, il y a peu de zones communes aux projections des aires V4 et V5.
L'intégration est une tache difficile qui nécessite un vaste réseau de connexions entre les 4 sytèmes spécialisés. Par exemple, pour comprendre un mouvement cohérent, le cerveau doit déterminer quels éléments du champ visuel se déplacent dans la même direction et à la même vitesse.
Les cellules du mouvement (aire V5) peuvent faire cette analyse car elles possèdent des champs récépteurs larges (plus larges que ceux des cellules de V1). Cependant, en raison même de la grande taille de leurs champs recépteurs, les cellules des aires spécialisées localisent mal les stimuli au sein du champ visuel. Pour obtenir une information spatiale précise, le cerveau doit se référer à une aire qui possède une carte précise de la rétine et du champ visuel. Ce sont les aires V1 et V2 qui possèdent les cartes les plus précises du champ visuel. Les aires spécialisées renvoient sans doute leurs informations vers ces aires où les résultats des comparaisons sont reportées sur le champ visuel.
Des recherches actuelles portent sur l'existence de connexions entre toutes les aires, qui permettraient une perception et une compréhension simultanées du monde (et non une intégration à un seul niveau, par regroupement des informations dans une seule aire maîtresse après que toutes les aires spécialiseés aient terminé leurs opérations).
Des connexions ont été montrées entre les aires spécialisées et les aives V1 et V2, de plus, les connexions "remontantes ou réciproques" (des aires spécialisées vers V1 et V2) sont organisées différemment des connexions "descendantes" (de V1 et V2 vers les aires spécialiseés). En effet, alors que seule une partie des aires V1 et V2 envoie des signaux aux différentes aires spécialisées, par exemple V5, les connexions remontantes de V5 se font vers toutes les cellules de la couche 4B de V1, même sur celles qui se connectent à V3 (ce qui peut permettre d'intégrer les informations de V5 concernant le mouvement avec celles de V3 concernant les formes tout en les localisant sur une carte précise du champ visuel).
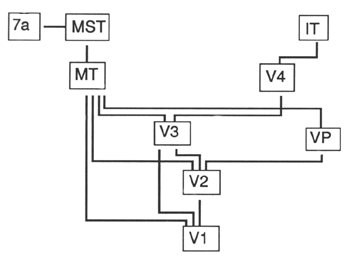 |
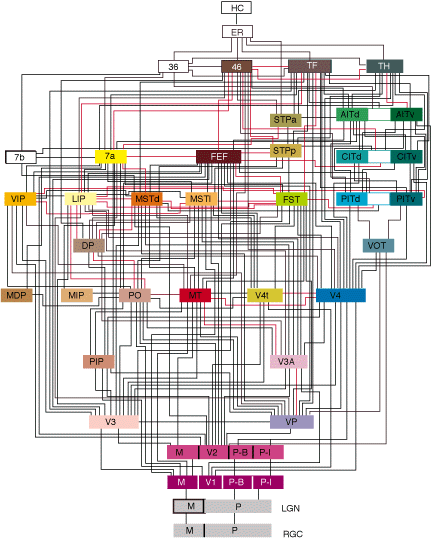 |
|
Figure 8. Relations entre les aires visuelles chez le singe Macaque (Van Essen) |
Figure 9. Relations entre différentes aires impliquées dans la vision chez le primate (Van Essen) |
 |
|
Figure 10. Triangle de Kanizsa Ce que V2 permet de voir: un triangle blanc. |
Dans cette illusion visuelle un observateur normal perçoit un triangle blanc (superposé à un triangle à contour noir) alors que le tracé de ce triangle n'est qu'ébauché. Il a été montré (Rudiger von der Heydt et Esther Peterhans) que les cellules sensibles à la forme de l'aire V1 ne réagissent pas lorsqu'on regarde la figure et ne détectent pas la ligne absente, mais les cellules de l'aire V2 reçoivent leurs informations de V1 et, avec leurs champs récepteurs plus larges et une fonction analytique supérieure, "déduisent" (construction visuelle) la présence d'une "ligne" (à la place du contour inexistant blanc). Il y a alors harmonisation des résultats des traitements effectués par les deux aires grâce aux connexions réciproques entre elles.