Glossaire
Histoire
Téléchargements
Hérédité des modalités de la division cellulaire et gène CDC2
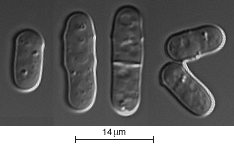
Dans un milieu favorable et une température de 35°, la levure
Schizosaccharomyces Pombe se divise toutes les 3 heures environ. Les cellules
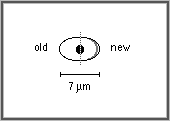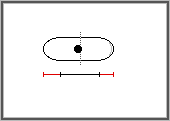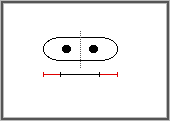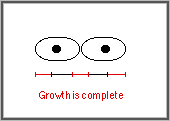
issues d'une division mesurent 6 à 7 micons ; elles grandissent
ensuite par leurs deux extrémités et entrent en division
lorsque leur taille atteint 13 à 14 microns. La division cellulaire
s'effectue par apparition d'une cloison au milieu de la cellule. L'entrée
en division en fonction de la taille est une caractéristique héréditaire.
 |
 |
Parmi les gènes en jeu, le gène CDC2 joue un rôle majeur : il code pour une protéine-kinase qui exerce un contrôle sur deux points crutiaux du cycle cellulaire, l'un est l'entrée en phase S, et l'autre est l'entrée en mitose. Le point de contrôle le plus important dans le cycle cellulaire de la levure fissipare est le point d'entrée en mitose. Le gène CDC2 est un composant décisif pour conduire les cellules en mitose ; lorsqu'il est défectueux, la mitose ne peut pas se produire. Les travaux sur le gène CDC2 et sur les autres gènes régulateurs du cycle cellulaire ont valu à P. Nurse, L Harwell et T. Hunt le prix Nobel de médecine 2001.
La séquence de l'allèle sauvage est désignée ci-après par cdc2sac-WT (Wild Type).
Les mutations de ce gène chez la levure Schizosaccharomyces Pombe se traduisent phénotypiquement par des changements dans la taille des levures. Toutes ces mutations sont des substitutions ponctuelles entraînant un changement ponctuel dans la séquence d'acides aminés de la protéine.
1 - Chez les mutants thermosensibles, la protéine codée
par ces allèles est fonctionnelle à température de
l'ordre de 25-26 degrés, mais est inactive à des températures
plus élevées, de l'ordre de 35 degrés. Ces souches
mutantes cultivées à 35 degrés n'entrent pas en mitose
et poursuivent leur croissance et atteignent ainsi une taille nettement
plus importante que celle de la souche sauvage.
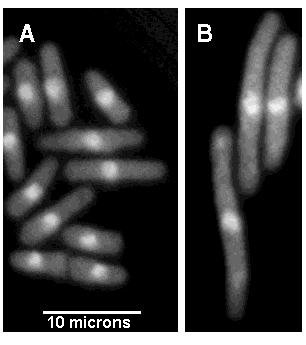
| Souche sauvage (A) et souche thermosensible (B) de S. Pombe cultivées
à 35°.
Les cellules ont été traitées par un colorant fluorescent qui révèle l'ADN du noyau. Les noyaux sont les sphères claires visibles au centre des cellules. Sur le clicé A on observe des levures de différentes tailles, donc à différents moments de leur croissance. La cellule la plus longue se trouve au bas du cliché A. Sa taille évaluée à partir de l'échelle fournie est de l'ordre de 13 microns, et on constate qu'elle achève sa division. Sur le cliché B, les cellules sont visualisées au même grossissement. Leur taille est de l'ordre de 17 à 18 microns. A la température de cette culture (35°) les cellules de cette souche mutante ne se divisent pas et poursuivent leur croissance jusqu'à atteindre une vingtaine de microns. En revanche elles se divisent à une température plus basse et les cellules obtenues ont le même comportement vis-à-vis de la température. |
 |
Parmi les nombreux allèles à l'origine de ce phénotype,
trois allèles sont retenus ici (TS pour thermosensible) :
| Nom de l'allèle | Nucléotide muté | Codon muté | Acide aminé muté |
| cdc2sac-TS1 | G124A | GAG42AAG | Glu42Lys |
| cdc2sac-TS2 | T628C | TTT210CTT | Phe210Leu |
| cdc2sac-TS3 | G634A | GGT212AGT | Gly212Ser |
2 - Chez d'autres mutants, en revanche, la protéine est hyperactive
et entraîne une entrée en mitose prématurée.
Les levures de ces souches ont toujours une taille nettement inférieure
à celle de la souche sauvage.
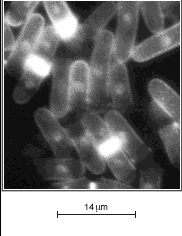
A |
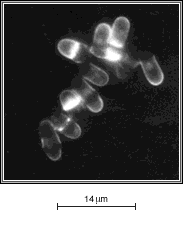
B |
Parmi les nombreux allèles à l'origine de ce phénotype,
deux allèles sont retenus (DP pour division prématurée)
:
| Nom de l'allèle | Nucléotide muté | Codon muté | Acide aminé muté |
| cdc2sac-DP1 | G200A | TGT67TAT | Cys67Tyr |
| cdc2sac-DP2 | G437A | GGC146GAC | Gly146Asp |
Unité du vivant et gène CDC2
Le gène CDC2 est un gène qui a été isolé et séquencé chez de nombreux organismes eucaryotes. La similitude globale entre le gène CDC2 de différents organismes eucaryotes et celui de la levure Schizosaccharomyces Pombe, est de l'ordre de 60 %. Cette similitude des séquences est indicatrice d'une homologie qui suggère l'idée que les mécanismes contrôlant le cycle cellulaire sont communs à tous les eucaryotes.
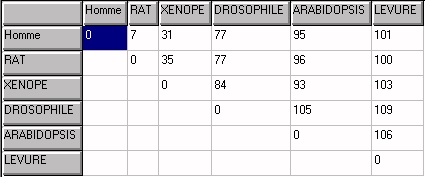
Tableau des distances
en valeurs absolues.
Alignement avec Clustalw. Matrice avec Phylogène.
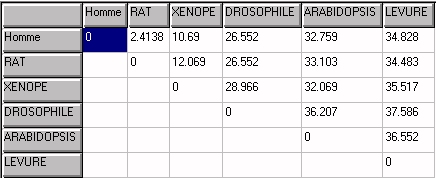
Tableau des distances
en pourcentage de la longueur.
Alignement avec Clustalw. Matrice avec Phylogène.
Cette unité du monde vivant est renforcée par le fait qu'un gène CDC2 d'une espèce, transféré dans une cellule d'une autre espèce, régit le cycle de la cellule hôte. Ainsi, des levures de la souche Schizosaccharomyces Pombe thermosensibles, dans lesquels on a transféré le gène CDC2 humain, deviennent capables de se diviser à des températures élevées.
Ce gène CDC2, commun à tous les eucaryotes, peut donc servir à argumenter l'origine commune de tous les eucaryotes.
Les gènes homéotiques servent de support pour développer l'idée que les mécanismes génétiques à la base de l'organisation antéro-postérieure sont partagés par tous les métazoaires.
Le gène CDC va plus loin dans la perception de l'unité
du vivant dans la mesure où il sensibilise à l'idée
que les mécanismes fondamentaux de la vie cellulaire sont partagés
par tous les organismes eucaryotes.