Glossaire
Histoire
Questions
Téléchargements
François Héron, Faculté de médecine, Caen
LA PROTEINE P 53
Le gène p53 est situé en position
sur le chromosome 17p13.1 Le gène a été très
conservé pendant l'évolution.
Structure biochimique
La protéine p53 est une phosphoprotéine
de 393 acides aminés de poids moléculaire 53 kDa. On la trouve
en très petite quantité dans les cellules normales, mais
en grande abondance dans les cellules transformées en culture ou
dans les tumeurs humaines.
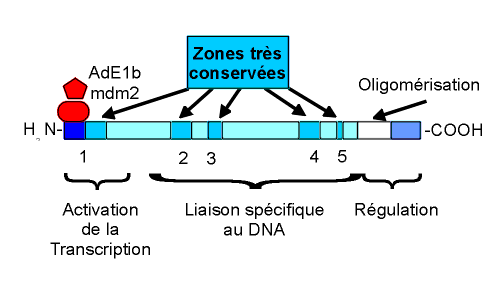 |
Schéma de la structure
biochimique de la protéine p53. Les segments indiqués par les flèches sont très conservés tout au long de lévolution. Dans la région amino-terminale, on note le site de lactivation de la transcription. Il est inhibée soit par la protéine mdm2 (augmentée dans certains cancers du sein) ou par la protéine AdE1b, des adénovirus pouvant provoquer des tumeurs expérimentales (inhibition de la fonction p53). En milieu de molécule, se trouve le site de liaison spécifique avec le DNA. Dans la région carboxy-terminale, se trouve le site favorisant la polymérisation qui est nécessaire à lactivité de reconnaissance du DNA. |
Pour être active, elle doit agir sous forme de tétramère. Aussi, une simple lésion sur une des protéines constitutives suffit à supprimer ou diminuer sa fonction.
La sensibilité à l'atteinte du DNA
Lors d'une agression cellulaire, la concentration et la demi-vie de p53 s'accroissent par une diminution de sa dégradation physiologique.
On a montré expérimentalement quune simple lésion double brin suffisait à induire une augmentation du taux de p53. De même, un trou denviron 30 à 40 bases dans la traduction du DNA sur un simple brin suffit également à provoquer cette augmentation et larrêt du cycle cellulaire. Un autre mécanisme déclenchant est constitué par un misappariement de bases ou une insertion délétion de bases.
Plusieurs mécanismes ont été suggérés pour expliquer larrêt de G1 provoqué par la protéine p53. Lactivation de la protéine p53 pourrait se faire par une liaison directe avec les lésions du DNA. Une autre mécanisme fait intervenir des protéine kinases activées par le DNA (DNA-PK), qui se fixent sur les liaison du DNA grâce à une protéine spécifique Ku, très conservée à travers lévolution.
Action de la P 53
Deux rôles particuliers sont dévolus à cette protéine : soit l'arrêt du cycle cellulaire entre la phase G1 et la phase S, soit la mort cellulaire par une phénomène d'apoptose.
La protéine p53 se lie avec une séquence spécifique de DNA (c'est donc un facteur de transcription) aboutissant à une interaction avec le cycle cellulaire par l'intermédiaire d'un gène appelé WAF1 / Cip1, (pour Wild Type p53-activated fragment et cdk2 inhibiting protein) dont la protéine p21 se lie aux kinases cdk2, et inhibe leur activité.
La cellule s'arrête avant la synthèse du DNA et peut réparer d'éventuels dommages.
De nombreux stimuli provoquent l'augmentation de la p53, notamment l'irradiation par les rayons X ou gamma.
Dans d'autres cellules, l'augmentation de la protéine p53 induite par l'irradiation provoque l'induction de l'apoptose ou mort programmée, que l'on peut interpréter comme la solution préférentielle pour l'organisme quand la réparation du DNA n'est pas possible.
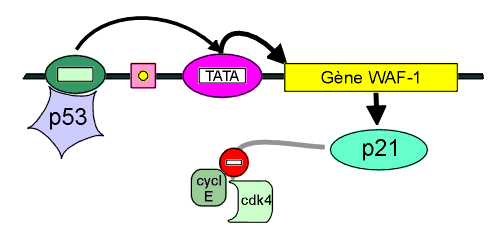
Schéma
de laction de la p53 comme facteur de transcription du Gène WAF-1,
nécessaire à la transcription de la protéine p21.
Celle-ci inhibe laction du couple
cycline E - cdk4 et empêche ainsi la phosphorylation de la protéine
chapeau Rb
(cf. plus loin). Il existe aussi une interruption de la formation dun
complexe
cycline E cdk2.
Lors de la radiothérapie ou la chimiothérapie qui attaquent le DNA, les cellules normales possédant la protéine sauvage p53 voient leur cycle cellulaire s'arrêter pour permettre une réparation du DNA (ou au pire une apoptose). Les cellules cancéreuses, qui ne possèdent pas le gène p53 'sauvage', vont progresser dans la synthèse du DNA, malgré l'agression, et aboutir à des cellules de plus en plus anormales qui finissent par mourir.
La protéïne p53, et le cancer
Dans de nombreuses pathologies cancéreuses, on observe des délétions ou des mutations de la protéine p53. Environ 60% des cancers ont une mutation de leur gène p53. Ces mutations sont du type sens inversé ou une homozygotie avec perte de l'allèle sauvage (cancers du colon, du poumon, de l'sophage, de l'estomac, du foie, du sein, de la vessie). Les mutations ont des localisations particulières à la pathologie cancéreuse mais reflètent aussi le cancérigène concerné (cf. le chapitre sur la cancérogenèse).