Glossaire
Histoire
Téléchargements
Rédigé par J-Y. Dupont, IA-IPR, Orléans-Tours
Ce point scientifique est fortement inspiré de :
- A. Stary, A. Sarasin : Xeroderma pigmentosum. La presse médicale n° 40, 20 décembre 1997).
- S. Doumer Queille : Rôle des ultraviolets dans la carcinogénèse cutanée. Thèse de l'université Paris XI, Avril 2001.
| Aux extrémités du spectre de la lumière visible, le rayonnement solaire comporte des infra-rouges et des ultra-violets. Le spectre UV est divisé en plusieurs régions : les UV de haute énergie (UVC) qui sont filtrés par la couche d'ozone, les UVB (qui représentent 0,3% du rayonnement parvenant à la surface de la Terre) et les UVA (0,1%).Toutefois, dans certaines régions du globe, la fragilisation de la "couche" d'ozone tend à élargir vers les UVC le spectre solaire parvenant à la surface de la terre . | 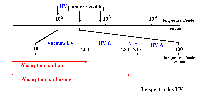
|
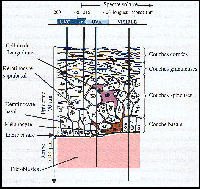
| Elle est constituée de l'épiderme
et du derme.
L'épiderme peut être subdivisé en quatre couches. Des kératinocytes indifférenciés prolifèrent au niveau de la lame basale et migrent vers les couches supérieures en subissant des modifications morphologiques et biochimiques. La différenciation terminale est la transformation en cellules cornées, mortes, en fome d'écailles qui constituent la barrière protectrice. D'autres types cellulaires sont présents dans l'épiderme : des mélanocytes, dans la couche basale, produisant les mélanines (protection contre les UV) et des cellules de Langerhans intervenant dans la réponse immune. Le derme est constitué de fibroblastes. Il renferme aussi des adipocytes impliqués dans la régulation de la température. |
|
Les UVC (haute énergie) pénètrent
faiblement dans la peau. Les UVB (5% des UV) pénètrent plus
profondément et atteignent la couche basale de l'épiderme.
Les UVA (9,5% des UV) pénètrent jusqu'au derme.
L'agression de la peau par les UV peut induire
des tumeurs cutanées :
- les mélanomes malins qui sont des tumeurs malignes très agressives et métastasiques provenant des mélanocytes. Bien que n'étant pas particulièrement localisés sur les zones corporelles exposées au soleil, l'exposition semble impliquée dans l'induction de ces mélanomes.
- les carcinomes cutanés qui peuvent avoir pour origine les cellules de la couche spineuse (carcinome spinocellulaire ou SCC) ou celles de la couche basale (carcinome basocellulaire ou BCC). L'âge moyen de la survenue est entre 50 et 60 ans. Les études épidémiologiques et expérimentales montrent que les carcinomes cutanés sont liés à l'exposition solaire : ils sont principalement localisés dans les zones photo-exposées ; il y a corrélation entre leur incidence et les faibles latitudes, la clarté de la peau, la déficience de la réparation des photolésions chez certains sujets ( cas du xeroderma pigmentosum).
ADN et UV
Les UV peuvent causer des dommages sur l'ADN,
les ARN et sur les protéines. Les effets majeurs sont cependant
les modifications produites directement par l'absorption des photons (émis
par les UVC et les UVB) par les bases de l'ADN.
| L'énergie absorbée
induit le plus souvent la formation d'une liaison covalente entre deux
pyrimidines adjacentes sur un même brin formant un dimère
de pyrimidine (DP).
On peut avoir 2 liaisons covalentes entre les C5 et les C6 de deux pyrimidines adjacentes. Cette dimérisation peut concerner les quatre types de séquences bipyrimidiques (TT,TC,CT,CC). Une liaison peut également s'établir entre les C6 et le C4 de deux pyrimidines adjacentes plus fréquemment sur des sites TC (quelquefois CC et TT). |
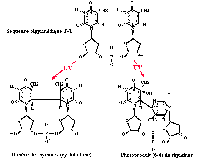
Cliquer pour agrandir |
Les dimères de pyrimidines ont pour conséquence une distorsion de la double hélice avec des répercussions sur la transcription, la réplication, la fixation des protéines qui se lient à l'ADN. Ces déformations sont de fortes courbures de l'ADN (7 à 44°).
Les mutations photo-induites
Certaines lésions de l'ADN peuvent bloquer la réplication. Toutefois certaines polymérases peuvent contourner la lésion. Dans ce cas , les mutations ponctuelles sont dues à l"incorporation d'une base incorrecte en face d'une lésion non réparée. La majorité des mutations sont des substitutions de bases : transition GC/AT, mutation en tandem CC vers TT.
Les cellules irradiées en phase G1 du cycle cellulaire ont le temps de réparer leur ADN si les lésions concernent le brin transcrit. Les mutations sont, dans ce cas, dues à la réplication des lésions situées sur le brin non transcrit.
Une irradiation en phase S ne laisse pas le temps
d'une réparation de l'ADN avant la réplication. Les mutations
résultent alors de la réplication des lésions situées
sur le brin transcrit.
Plusieurs processus permettent de détecter et réparer les lésions induites sur l'ADN.
1 - La photo-réactivation
Elle est induite par la lumière et est
catalysée par des ADN photolyases : l'enzyme provoque la cassure
des liaisons C5-C6, C6-C6 ou C4-C6.
2 - La réparation par excision de bases
(BER)
Elle consiste en un retrait des bases modifiées
lorsque celles-ci n'entraînent pas de distorsion de la double hélice.
Les bases modifiées sont purement et simplement enlevées
par des glycosylases et des endonucléases.
3 - La réparation par excision de nucléotides
(NER)
Elle est impliquée dans le retrait de
lésions volumineuses qui provoquent une distorsion de la double
hélice. Il existe deux voies de réparation :
- la réparation couplée à la transcription qui concerne la réparation rapide des lésions situées sur le brin transcrit et affectant des gènes en cours de transcription.
- la réparation globale du génome qui concerne la réparation lente de l'ADN pour des gènes qui ne sont pas en cours de transcription.
| La NER, chez les Mammifères,
se déroule en 4 étapes :
1 - Recherche et reconnaissance de la lésion 2 - Ouverture de la double hélice au niveau de la lésion 3 - Incision de part et d'autre de la lésion puis excision pour éliminer le fragment d'ADN portant la lésion 4 - Resynthèse puis ligation. Chez l'Homme, une trentaine de protéines sont impliquées. Seules certaines d'entre elles seront envisagées ici. |
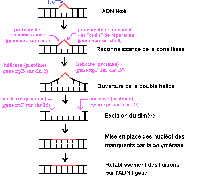
|
- La recherche et la reconnaissance de la lésion
- L'ouverture de la double hélice
- L'incision de part et d'autre de la lésion et l' excision
- La resynthèse puis ligation
Dans le cas d'une réparation couplée à la transcription, c'est le blocage de l'ARN-polymérase au niveau de la lésion qui permettrait de recruter des protéines de couplage transcription-réparation (CS-A et CS-B) et déclencherait ainsi les mécanismes de réparation. Dans le génome inactif, c'est la protéine xpC associée à d'autres protéines qui assurerait ce rôle de recherche des lésions.
Le xeroderma pigmentosum (=XP)
C'est une affection génétique rare transmise sur le mode autosomal récessif avec une fréquence d'environ 1/106 en Europe et aux Etats Unis, de 1/105 au Japon et en Egypte. Dans ces derniers cas, 30% des parents de sujets atteints de XP sont consanguins et 21% cousins au premier degré.
Principales
caractéristiques cliniques
|
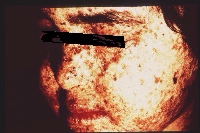
|
Les caractéristiques
cellulaires
| Les cellules cultivées in vitro provenant de malades
XP ont, après irradiation aux UVC, une fréquence de mutations
beaucoup plus importante que les cellules normales soumises aux mêmes
agents. La présence de lésions non réparées
provoque, lors de la réplication de l'ADN endommagé, l'insertion
de bases anormales en face des lésions et induisent ainsi des mutations
dans le génome des cellules filles.
La fréquence particulièrement importante de cancers cutanés dans les zones exposées au soleil est corrélée avec le taux élevé de mutations détectées sur plusieurs loci. |
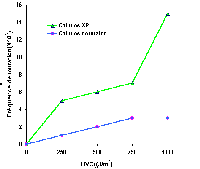
|
Le spectre de mutation particulièrement caractéristique de l'action des UVC (transition C vers T, double mutation CC vers TT) est retrouvé sur les oncogènes ras activés et sur le gène suppresseur p53 dans les cellules tumorales des patienst XP.
Les caractéristiques génétiques et biochimiques
Dans les cellules XP, une des premières étapes de la réparation
par excision-resynthèse (NER) est déficiente. La plupart
des gènes de réparation impliqués ont été
identifiés : les méthodes ont reposé le plus souvent
sur l'introduction dans des cellules sensibles aux UV de fragments d'ADN
restaurant chez ces cellules un comportement normal. Le clonage et séquençage
de ces gènes ont permis de connaitre la séquence en acides
aminés des protéines impliquées et ainsi de déduire
la fonction de ces enzymes.
Au moins huit gènes seraient impliqués dans le syndrome
XP.
| L'UDS (Unscheduled DNA
Synthesis) est la méthode biologique la plus fréquemment
utilisée.
Des cellules préalablement irradiées aux UV sont mises en présence de thymidine tritiée. Une cellule normale incorporera d'autant plus de ce précurseur radioactif pour réparer son ADN que la dose d'UV reçue aura été plus importante. Une cellule XP, incapable de réparer correctement les dommages, n'incorporera que peu de précursseur marqué. La mesure est effectuée par autoradiographie en comptant le nombre de grains d'argent par noyau. |
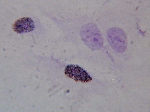
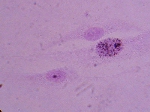
Cliquer pour agrandir |
| Cette méthode est appliquée sur des cellules foetales cultivées in vitro provenant de biopsie de trophoblaste ou de liquide amniotique. L'efficacité de réparation de l'ADN est comparé à celle de cellules issues de biopsie de peau des parents normaux. |
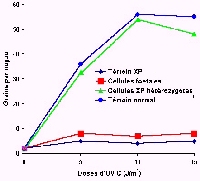
Cliquer pour agrandir |
| La prévention en mettant l'individu à
l'abri de la lumière reste la meilleure solution thérapeutique.
On pratique l'ablation chirurgicale de tumeurs et l'utilisation de composés rétinoïdes pour empêcher la progression tumorale. On utilise également l'autogreffe d'une partie de la peau non exposée au soleil. L'identification des gènes impliqués a conduit à établir des protocoles de thérapie génique cellulaire : la culture in vitro de kératinocytes de patients atteints de XP permet de réaliser la "reversion phénotypique" de ces cellules par transfert des gènes déficients de réparation de l'ADN à l'aide de vecteurs retroviraux. |
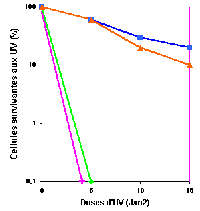
Cliquer pour agrandir |