Rôles connus des mitochondies dans la reproduction
Rédigé par Françoise Jauzein (INRP/ENS-Lyon),
à partir de "Mitochondries et reproduction",
un article de Pascale May-Panloup, Marie-Françoise Chrétien,
Yves Malthièry et Pascal Reynier,
paru dans Médecine/Sciences 2004; 20:779-83
Deux des caractéristiques essentielles des mitochondries sont de
jouer un rôle central dans le métabolisme énergétique
cellulaire et de posséder leur propre génome, transmis de
façon maternelle, par le cytoplasme de l'ovocyte, au moment de la
fécondation. Le fait qu'elles puissent être impliquées
dans les problèmes liés à la reproduction humaine
est une idée relativement récente, liée en partie
au fait que, malgré des avancées techniques continues en
matière d'assistance médicale à la procréation,
le taux moyen de grossesses obtenues après fécondation in
vitro (FIV) ne dépasse pas 25% par transfert d'embryon.
Les échecs de la fécondation ou du développement
embryonnaire peuvent être mis en relation avec la qualité
et la maturation des gamètes, dans lesquels les mitochondries constituent
un facteur clé.
La mitochondrie : des rôles multiples et son propre ADN
Les mitochondries jouent non seulement un rôle fondamental
dans le catabolisme oxydatif aboutissant à la production d'une forme
utilisable d'énergie, l'ATP, mais elles jouent également
un rôle déterminant dans l'apoptose, la thermogénèse,
l'homéostasie du calcium, et de nombreuses voies anaboliques comme
la synthèse de l'hème, des protéines fer-soufre, des
nucléotides ou encore des stéroïdes.
Les proteines mitochondriales humaines sont au nombre de mille environ.
Ce protéome est codé pour l'essentiel par le génome
du noyau . Le génome mitochondrial (ADN circulaire double brin)
est assez petit (16 569 paires de bases) et se compose notamment de gènes
d'ARN ribosomiques, d'ARN de transfert, et de protéines de la chaîne
respiratoire.
Dans une cellule humaine, le nombre de copies d'ADN mitochondrial (ADNmt)
varie entre quelques dizaines et plusieurs milliers, selon le type cellulaire.
Le fait de posséder une séquence unique de la molécule
d'ADNmt dans toutes ses cellules est appelé homoplasmie et
constitue la règle chez la plupart des êtres vivants. L'hétéroplasmie
correspond à la présence dans une cellule ou un tissu de
deux types de génomes mitochondriaux différents par des mutations
ou du polymorphisme génique.
I Implication des mitochondries dans la fertilité
A. Fertilité masculine
Dans les spermatozoides humains, les mitochondries, très
différenciées, constituent 11 à 13 tours de
deux mitochondries par tour, au niveau de la pièce intermédiaire.
Au cours de la spermatogénèse, s'est produite une réduction
importante du nombre de mitochondries par cellule et de copies du génome
mitochondrial (arrêt de la réplication de l'ADNmt).
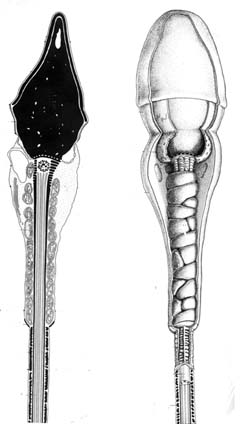 |
Reconstitution du spermatozoide à partir de vues au microscope
électronique
http://spermiologie.u-strasbg.fr |
Fertilité masculine et ATP
Le pouvoir fécondant du sperme est directement lié à
l'activité énergétiques de ces mitochondries (elles
délivrent à l'axonème l'ATP indipensable au mouvement
du flagelle). L'incubation de sperme de patients infertiles en présence
d'ATP améliore le taux de fécondation in vitro.
Cependant les mitochondries paternelles ne particient pas au développement
embryonnaire. Le traitement par le cyanure (bloquant de la chaîne
respiratoire) des spermatozoides injectés dans un ovocyte, ne perturbe
pas le développement de l'oeuf.
Fertilité masculine et variation quantitative de l'ADNmt
Le nombre moyen de copies d'ADNmt présentes dans des spermatozoides
matures peut être estimé grâce à des techniques
de PCR, chez la souris il est de 10 en moyenne par spermatozoide (15 fois
moins que dans les spermatides).
Chez l'homme, le nombre de copies d'ADNmt estsignificativement
augmenté dans les spermatozoides anormaux; alors que les spermatozoides
de bonne qualité en sont quasiment dépourvus.
On peut donc considérer le taux d'ADNmt
comme un bon marqueur de la maturation des spermatozoides.
Taux moyens d'ADNmt par spermatozoide
dans deux fractions spermatiques obtenues
pour chaque patient,
après séparation par gradient
de densité:
fraction de forte densité correspondant
aux spermatozoides normaux (fécondants)
et de faible densité aux spermatozoides
anormaux.
|
Nombre de copies d'ADNmt par spermatozoide
|
|
Fraction de forte densité: spermatozoides normaux
|
0 à 3
|
moyenne :1
|
Fraction de faible densité:
spermatozoides anormaux
|
1 à 22
|
moyenne: 12
|
Mesure par PCR quantitative en temps réel.
Résultats compilés sur 31 patients.
Fertilité masculine et variation qualitative de l'ADNmt
Différentes observations tendent à établir un lien
entre fertilité et anomalies qualitatives de l'ADNmt.
Une oligoasthénospermie a été obervée
chez des patients présentant des mutations ponctuelles ou des délétions
de l'ADNmt.
Certains haplogroupes mitochondriaux ont été corrélés
à des qualités spermatiques moindres (en particulier l'haplogroupe
T , surreprésenté chez les hommes asthénospermiques,
et qui se caractérise par une baisse d'activité de certains
éléments de la chaîne respiratoire).
B. Fertilité féminine
Au cours de l'ovogenèse, le cytoplasme de l'ovocyte subit une croissance
phénoménale (son diamètre passe de 30 à 120
micromètres): il accumule un grand nombre de substances (transcrits,
substrats méaboliques...) et son contingent de mitochondries évolue
beaucoup: il subit d'abord une restriction puis une intense amplification.
Les ovogonies ne contiennent qu'une dizaine de copies d'ADNmt alors
que les ovocytes mûrs en renferment jusqu'à plusieurs dizaines
de milliers. Il se produit une expansion clonale à partir d'un très
petit nombre d'ADNmt selectionnés, ce qui permet à l'ovocyte
de transmettre au nouvel individu une population homoplasmique d'ADNmt.
Ce "goulot génétique" permet normalement d'éliminer
les rares formes mutées, sauf dans le cas où la sélection
inclut un ADNmt muté. On observera alors, une accéleration
d'une éventuelle dérive génétique, avec fixation
d'une mutation dans la descendance (fort utile pour
retrouver, parmi les "Eves" fondatrices européennes, la lignée
maternelle dont chacun de nous est issu).
Fertilité féminine et variation quantitative de l'ADNmt
Dans les ovocytes humains (obtenus en vue de FIV), on trouve entre 50 000
et 100 000 copies d'ADNmt, mais une très grande variabilité
existe entre ovocytes d'une même femme, y compris entre ceux d'une
même cohorte.
Néanmoins, il est maintenant établi que les cohortes
d'ovocytes présentant des échecs de fécondation in
vitro sont significativement moins riches en ADNmt que ceux qui présentent
un taux normal de fécondation. Dans l'espèce humaine il existe
une relation nette entre la richesse d'un ovocyte en ADNmt et sa fécondabilité;
le taux d'ADNmt pouvant ainsi être considéré (tout
comme pour le spermatozoide, mais en sens inverse) comme un bon marqueur
de maturation cytoplasmique.
Taux moyens d'ADNmt dans des ovocytes isolés après
échec de FIV.
Groupe A: échec de fécondation pour cause spermatique
vraisemblable (ovocytes normalement fécondables)
Groupe B: échec de fécondation sans cause reconnue
(ovocytes de mauvaise fécondabilité)
|
Nombre de copies d'ADNmt pour1000 ovocytes
|
Groupe A :
ovocytes normaux
|
de 210 à 310
|
moyenne: 250
|
Groupe B :
ovocytes de mauvaise qualité
|
de 90 à 200
|
moyenne: 130
|
Mesure par PCR qantitative en temps réel
Résultats compilés, pour le groupe A sur 21 patientes,
pour le groupe B sur 47 patientes.
Fertilité féminine et variation qualitative de l'ADNmt
Il existe peu de cas connus de mutation de l'ADNmt ovocytaire. La plus
courante est une délétion d'environ 5000 paires de bases,
retrouvée dans 33 à 66% des ovocytes humains. Le taux d'hétéroplasmie
moyen de ces délétions est de 0,1%, ce qui n'a vraisemblablement
pas de retentissement fonctionnel.
Cependant, de récentes données sur les ovocytes bovins,
montrent que certains haplogroupes mitochondriaux influencent nettement
le taux de production de blastocystes.
II Implication des mitochondries dans le développement embryonnaire
A. La situation naturelle
ATP et développement embryonnaire
Après la fécondation, seules les mitochondries apportées
par l'ovocyte sont conservées et serviront de "réserve" initiale
des mitochondries du nouvel individu.
Ces mitochondries d'origine maternelle, initialement dispersées
dans le cytoplasme de l'ovocyte, sont redistribuées dans le zygote,
autour des deux pronuclei (vraisemblablement en relation avec la consommation
d'énergie due aux évènements nucléaires).
Puisque l'inhibition de la transcription et de la traduction mitochondriale
ne perturbe pas le développement du zygote jusqu'au stade blastocyste
(chez la souris), on peut supposer que le contingent mitochondrial maternel
suffit à assurer les besoins énergétiques de l'embryon
jusqu'à ce stade.
Par la suite, à un stade qui varie selon l'espèce, on
assiste chez l'embryon, à une forte augmentation de la production
d'ATP, en parallèle avec une augmentation des protéines de
la chaîne repiratoire, de profonds remaniements morphologiques des
mitochondries (décondensation de la matrice mitochondriale, élongation
des mitchondries et augmentation du nombre de crêtes) et une élévation
de la consommation de substrats énergétiques.Cependant, il
ne semble pas y avoir de réplication de l'ADNmt avant l'implantation
de l'embryon. La plupart des études menées chez la souris
montrent un taux d'ADNmt constant jusqu'à la gastrulation.
Comme le montre la baisse de la capacité de développement
des embryons dans le cas d'une réduction expérimentale du
taux d'ATP ovocytaire (chez la souris), le contenu en ATP constitue un
bon indicateur de la vitalité de l'embryon.
Il est possible en effet, de "sauver" (chez l'animal) des ovocytes
de mauvaises qualités par transfert de cytoplame provenant d'un
ovocyte normal, ou de façon plus spécifique, par transfert
de mitochondries purifiées, capable à lui seul d'augmenter
la production d'ATP et de prévenir l'apoptose de l'ovocyte receveur.
Devenir des mitochondries d'origine paternelle
La transmission uniparentale de l'ADNmt, classiquement admise, implique
une destruction totale de l'ADNmt d'origine spermatique dans l'oeuf fécondé.
L'élimination spécifique des mitochondries paternelles dans
l'ovocyte et l'absence de réplication de l'ADNmt au sein de l'ovocyte
fécondé sont les mécanismes qui permettent d'expliquer
l'absence de transmission d'ADNmt paternel.
La reconnaissance et la destruction des mitochondries paternelles reposent
sur le processus d'ubiquitinylation d'une protéine membranaire mitochondriale
(la prohibitine), commencé au cours de la spermatogenèse.
Lors de la décondensation du spermatozoide dans le cytoplasme ovocytaire,
les sites ubiquitinylés des membranes mitochondriales paternelles
(sites initialement masqués par des ponts disulfures durant le transit
épididymaire) seraient alors démasqués et pris pour
cible par les enzymes protéolytiques du cytoplasme ovocytaire.
Cette destruction se produirait au plus tard lors de la troisième
division de segmentation de l'embryon.
Ce mécanisme semble "spécifique" au sens propre du terme,
car dans des embryons "chimères", issus de croisements murins interspécifiques
(entre espèces de souris différentes), on observe une persistance
des mitochondries paternelles. Ce mécanisme est également
spécifique des mitochondries de la lignée germinale mâle,
puisque des mitochondries d'origine tissulaire différente (de foie
par exemple) injectées dans un ovocyte de souris ne sont pas détruites,
et que l'on retrouve de l'ADNmt "étranger" chez les nouveaux-nés.
B. Les situations d'assistance médicale à la procréation
Hétéroplasmie naturellement possible
Il semble que le dogme de l'uniparentalité des mitochondries soit
remis en cause dans un certain nombre de cas (très faible cependant).
Il a été possible de retrouver, chez l'homme, de l'ADNmt
d'origine paternelle dans des tissus extraembryonnaires, de même
que dans les muscles d'un patient atteint d'une forme mitochondriale de
myopathie.
Hétéroplasmie conséquence d'AMP
Certaines techniques d'AMP comme le clonage ou le transfert de cytoplasme,
fournissent à l'ovocyte receveur un volume important de cytolasme
exogène, et apportent donc un nombre non négligeable de mitochondries
de la même espèce mais qui, n'étant pas d'origine paternelle,
ne seront pas éliminées par l'ovocyte receveur. Il s'en suivra
un individu à cytoplasme hétéroplasme, ce qui n'est
pas sans poser un certains nombre de problèmes éthiques.
On peut remédier aux échecs de fécondation chez
des patientes présentant une mauvaise qualité ovocytaire,
en réalisant, en même temps que l'ICSI (avec le sperme du
conjoint), une injection de cytoplasme d'un ovocyte donneur de bonne qualité
(provenant d'une autre femme), à raison de 5 à 15% du volume
de l'ovocyte receveur. Une trentaine d'enfant, principalement aux Etats-Unis,
ont ainsi déjà vus le jour. L'hétéroplasmie
se retrouve après la naissance, dans des proportions variables selon
les tissus.
Dans le cas de clonage embryonnaire ou somatique (le clonage humain
reproductif est toujours interdit, mais le clonage humain à but
thérapeutique est déjà autorisé en Angleterre
par exemple), on va faire fusionner (par electroporation) une cellule entière
(embryonnaire ou somatique), avec un ovocyte énucléé.
Certes, le volume cytolasmique de l'ovocyte est toujours bien supérieur
à celui de la cellule (notamment s'il s'agit de cellule différenciée),
mais ce dernier n'est pas negligeable.
Une des hypothèses avancées par les spécialistes
pour expliquer le faible taux de réussite du clonage animal (3%
au maximum), quelque soit la technique utilisée (cellule donneuse
embryonnaire ou somatique, en phase G1 ou G0), est l'hétéroplasmie
de l'oeuf obtenu dans ces conditions.
Dans le cas du clonage thérapeutique chez l'homme, toutes les
cellules souches (cellules ES) issues de l'embryon obtenu par fusion entre
un ovocyte humain énucléé et une cellule somatique
du patient, devraient présenter une forte héteroplasmie.
Outre les problèmes éthiques que posent la création
d'embryon ou de tissus porteurs d'information génétique multiple
(trois "parents" dans le cas d'une ICSI avec transfert de cytoplasme),
il est impossible de prévoir actuellement les conséquences
à long terme, et en particulier les éventuels risques pathogènes,
de telles hétéroplasmies.
Glossaire spécifique
Haplogroupes mitochondriaux: combinaisons spécifiques
de polymorphismes géniques reflétant l'évolution des
populations
Ubiquitinylation: marquage de protéines endogènes
leur permettant d'être adressées vers le protéasome
qui les dégradera en peptide