Glossaire
Les innovations génétiques à l'origine de la résistance des moustiques aux insecticides OP et aux carbamates
- Les innovations génétiques à l'origine d'une production accrue d'estérases
- Les innovations génétiques à l'origine d'une ACE insensible aux insecticides
a ) Génome des moustiques sensibles
-
On connaît deux gènes, dont les loci sont situés sur
le même chromosome et espacés de 2 à 6 kb, qui codent
pour des estérases : Est 3 (codant pour l'estérase A) et Est2
(codant pour l'estérase B). Il existe un polymorphisme important pour
chacun de ces gènes. Ce polymorphisme est neutre, c'est à dire
sans conséquences sur l'activité enzymatique.
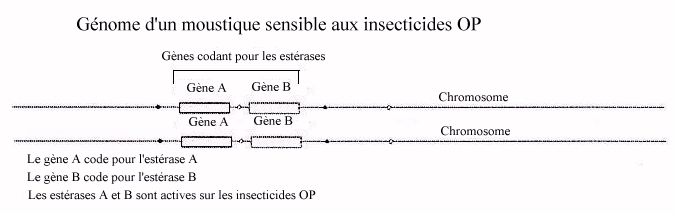
- Les estérases A et B hydrolysent toutes les deux des liaisons ester, mais avec une spécificité relative à la position des liaisons (dans le test au papier filtre présenté dans ce dossier, le substrat chromogène est double : alpha naphtylacetate et beta naphtylacetate, et est donc hydrolysé spécifiquement par l'estérase A pour le premier et par l'estérase B pour le deuxième) Dans un souci de simplification, ces gènes ont été désignés par les lettres A et B dans la banque de données fournies, et les allèles de chacun de ces gènes sont désignés par des numéros (A1, A2, ... B2, B4, ....).
- Les moustiques dont la résistance est due à une production accrue d'estérases ont un génome caractérisé par une amplification des gènes Est3 (ou gène A) et Est2 (ou gène B) : chacun de ces gènes est présent en de nombreux exemplaires. L'innovation génétique consiste donc en duplications géniques. Tous les gènes résultant de ces duplications sont identiques et s'expriment, ce qui explique la production accrue d'estérases. On désigne par "ester" le "supergène" résultant de l'amplification (ester B pour le résultat de l'amplification du gène B, ester A-B pour le résultat de l'amplification de l'ensemble des deux gènes A et B).
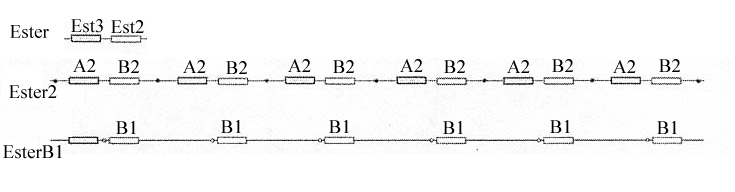
Selon l'allèle du gène qui est amplifié, on distingue plusieurs "supergènes ester" : ester B1 (plusieurs copies de l'allèle B1), ester A2-B2 (plusieurs copies de l'ensemble des deux allèles A2-B2), ....
- L'existence de plusieurs "supergènes ester" différents (ester B1, ester B4, ester A2-B2, ester A4-B4, ....) indique que plusieurs innovations génétiques à l'origine de la résistance sont intervenues de façon indépendante dans les populations de moustiques (duplications à partir de l'allèle B1, duplication à partir de l'allèle B4, ...). Cependant, le polymorphisme des gènes A et B est très nettement supérieur à celui des "supergènes ester" ; cela indique donc que les mutations à l'origine des "supergènes ester" sont relativement rares. D'autre part, on a pu constater que des allèles ester apparus en premier dans une région, se retrouvaient ensuite dans une autre région assez éloignée ; cela s'interprète comme le résultat de migrations des moustiques (par transport aérien ou maritime par exemple) et non comme le résultat de deux mutations identiques indépendantes. Le niveau d'amplification varie selon les allèles ester : il peut atteindre 100 copies du gène ou plus pour ester B1, tandis qu'il ne dépasse pas quelques copies pour ester A4-B4. De plus, pour un même allèle (exemple ester B1) le niveau d'amplification peut être variable selon les individus au sein d'une population sauvage ou entre des populations différentes.
- Enfin, il existe un allèle ester A1 pour lequel la production accrue d'estérases n'est pas due à l'amplification du gène A1, mais à un changement dans la régulation de l'expression de ce gène. Comme les processus de régulation génique ne sont pas au programme de Tle S, nous n'avons pas retenu cet allèle dans la banque de données.
-
C'est en 1978 que sont apparus dans la région de Montpellier des
moustiques résistants aux insecticides OP, dont la résistance
est due à la production d'une AchE insensible. Ces moustiques sont
homozygotes pour un allèle mutant du gène qui code pour l'AchE.
Les séquences des allèles AchES (allèle présent
chez les moustiques sensibles) et AchER (allèle mutant présent
chez les moustiques résistants) viennent d'être publiées
(Nature, Mai 2003) . Leur comparaison révèle 28 substitutions
dont 27 sont muettes (aucune influence sur la séquence polypeptidique)
et une est à l'origine du changement d'un acide aminé (substitution
G/A en position 739 ; glycine 247 de l'AchE sensible remplacée par
une sérine dans l'AchE résistante). Cet acide aminé
est situé dans le site actif, près du site catalytique. Des
expériences de transgenèse (intégration de l'allèle
AchER dans des cellules de drosophile) ont permis de confirmer l'insensibilité
de l'AchE codée par cet allèle AchER.
- En 1993, dans la région de Montpellier, est apparu un nouvel "allèle" appelé AchERS (appelé AchE 93 dans les données fournies sur ce site) ; il s'agit en fait d'un "supergène" formé de deux copies en tandem du gène AchE : l'une de ces copies est l'allèle AchES et l'autre AchER. Cette innovation génétique résulte donc d'une duplication génique. L'interprétation classique suggérée par le programme et utilisée pour les familles multigéniques est la suivante : il y a eu duplication de l'allèle AchES, suivie de mutations d'une des copies conduisant à l'allèle AchER. Cependant, l'interprétation qui a les faveurs des chercheurs est différente : chez un individu hétérozygote AchES/AchER, il y a eu au cours de la méiose un crossing-over inégal aboutissant à réunir sur un même chromosome les deux allèles AchER et AchES. Un seul événement génétique, une duplication génique, serait donc à l'origine de ce "supergène". Ces moustiques produisent donc les deux types d'AchE, comme les hétérozygotes AchER/AchES, mais cette capacité est transmise de génération en génération à cause de la liaison très étroite entre les loci des gènes AchER et AchES.
Une mutation identique a été identifiée chez une autre espèce, l'anophèle.