Glossaire
Histoire
Téléchargements
Détermination hiérarchisée du sexe chez les mammifères
CHEZ LES MAMMIFERES
Rédigée par Françoise Jauzein, Lycée
Berthollet, Annecy
Relue par Corinne Cotinot, INRA
Depuis les travaux de Jost, dans les années 40, on sait que l'ablation des gonades d'un foetus de mammifère sexuellement indifférencié entraîne un développement femelle, aussi bien au niveau du tractus interne que des organes génitaux externes, et ceci, quelque soit son sexe génétique. Ainsi, la détermination du sexe est équivalente à la détermination des testicules.
Le fait que la détermination des testicules dépende de
la présence d'un chromosome Y est connu depuis le début des
années 60, et a été déduit de l'observation
d'anomalies chromosomiques: des humains 45,XO (Turner) ou des souris XO
ne montrent pas de développement de tissu testiculaire. De plus,
un caryotype humain 47,XXY (Klinefelter) entraîne un phénotype
mâle. Ceci prouve que, chez les mammifères, la détermination
des testicules dépend de la présence d'un chromosome Y et
non du nombre de chromosome X.
Le facteur, encore inconnu, codé par un gène du chromosome
Y, et déterminant la différenciation des testicules, a été
appelé TDF (testis determining factor).
LE DETERMINANT TESTICULAIRE : SRY
L'identification de TDF a été réalisée en
1989, grâce à l'étude d'individus humains dont le sexe
phénotypique est en discordance avec le sexe chromosomique; hommes
possédant deux X (46,XX) ou femmes possédant un X et un Y
(46,XY).
Ces individus phénotypiquement mâles et porteurs d'un
caryotype 46,XX, renfermaient dans leur génôme une séquence
provenant du chromosome Y. L'analyse de ce segment
d'ADN montra une séquence codante (open-reading frame ou ORF) qui
fut appelée: "région déterminant le sexe" du chromosome
Y ou SRY (Sex-determining Region of Y chromosome).
Ce gène est situé sur le bras court du chromosome Y humain
et répond aux caractéristiques attendues d'un "TDF":
-ce gène est conservé sur le chromosome Y de nombreuses espèces de mammifères;
- la protéine déduite du gène SRY possède un domaine permettant sa liaison à l'ADN. Ce domaine, homologue à celui des protéines du HMG (high mobility group) définit une nouvelle famille de gènes appelé SOX (SRY-related HMG-box) qui interviennent dans de nombreux processus de développement;
-la chronologie de l'expression de ce gène coïncide avec la période de détermination du sexe, il s'exprime chez la souris de 10,5 à 12 jours post coïtum, spécifiquement dans les cellules somatiques de la crête génitale mâle, ce qui correspond aux jours qui précèdent la détermination testiculaire. On trouve des transcrits (ARNm) de ce gène dans le testicule adulte murin et humain;
-des individus qui développent un phénotype femelle malgré la présence d'un Y (46,XY) présentent une mutation dans le gène SRY, le rendant non fonctionnel;
-enfin la meilleure démonstration fut obtenue par transgénèse chez la souris; l'introduction d'un fragment de 14Kb d'ADN contenant le gène Sry (en minuscule = murin), dans une souris femelle XX, entraîne le développement de testicules, ce qui suggère que c'est le seul gène du chromosome Y impliqué dans la détermination testiculaire.
SRY est donc responsable de l'établissement du sexe gonadique, c'est le TDF tant attendu!
Période d'expression et mode d'action de SRY
Chez la souris, la transcription de Sry pendant la vie foetale est limitée
à la période de formation des crêtes génitales
entre 10,5 et 12,5 jpc (jours post coïtum). Les premières cellules
dans lesquelles ce gène s'exprime sont les précurseurs des
cellules de Sertoli. Ceci permet, juste avant la formation des cordons
séminifères, d'entraîner la différenciation
des cellules de soutien vers la voie mâle.
Chez les autres mammifères, dont l'homme, le gène SRY
est transcrit pendant une période plus longue dans les cellules
de Sertoli, de la formation des crêtes génitales jusqu'après
la différenciation de ces cellules. Dans le testicule adulte, ce
gène s'exprime dans les cellules germinales méiotiques et
post-méiotiques.
 |
|
(Demande de droits en cours) |
La protéine SRY humaine se compose de 204 acides aminés.
Elle se lie spécifiquement, par son domaine HMG renfermant environ
80 acides aminés, à l'ADN bicaténaire portant la séquence
5'A/TAACAAA/T 3'. Après fixation de cette protéine, la molécule
d'ADN subit une courbure de 70 à 80° dans la direction du grand
sillon. Cela pourrait permettre le rapprochement et l'interaction entre
des facteurs de transcription fixés sur le ou les gènes cibles
de SRY, dont les sites de liaison sont éloignés les uns des
autres.
Comme on retrouve la protéine SRY dans le noyau des cellules
de Sertoli mais aussi dans les cellules germinales, on peut penser que
ce gène permet non seulement la différenciation de la gonade
vers la voie mâle mais présente aussi d'autres fonctions.
Ainsi le sexe génétique
est déterminé à la fécondation, selon
le chromosome sexuel apporté par le spermatozoïde. L'établissement
du sexe gonadique est sous contrôle génétique
et dépend de la présence du gène SRY.
Suite à l'action du gène SRY, une cascade d'autre
gènes est activée dans le testicule. Tandis qu'en l'absence
de SRY et en présence de deux chromosomes X, la gonade se différencie
en ovaire. Cette période de différenciation gonadique se
réalise chez l'homme entre la 5ème et 8ème semaine
de gestation.
L'établissement du sexe phénotypique
se réalise, quant à lui, sous l'effet des hormones
sécrétées par la gonade
différenciée, à deux périodes de la
vie: le tractus génital se différencie
pendant la vie foetale et les caractères sexuels secondaires, à
la puberté.
LES AUTRES GENES DE LA CASCADE
|
|
(humaine) |
|
(souris) |
(humaine) |
|
| SRY | bras court du chr Y | précurseurs et cellules de Sertoli, cellules germinales mâles | juste avant la différenciation des testicules
reprise à l'âge adulte |
204 aa, possède un domaine de liaison à l'ADN entraînant sa courbure |
|
|
|
|
cellules de Sertoli ou cellules de la granulosa, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Le gène WT1
Ce gène, localisé sur le bras long du chromosome 11, mesure
50kb et comprend 10 exons. Il code pour différentes isoformes protéiques
(selon l'épissage et le processus d'édition de l'ARN), dont
un facteur de transcription se liant à l'ADN par des domaines en
doigt de Zinc.
Il s'exprime très tôt dans le développement embryonnaire
des souris, dès le 9ème jour post-coïtum, au niveau
de la crête urogénitale, puis toute la vie dans les gonades,
au niveau des cellules de Sertoli du testicule et de la granulosa de l'ovaire.
Il existe différents phénotypes sexuels associés à des mutations du gène WT1. Le syndrome de Denys-Drash qui touche des enfants en bas âge, des deux sexes, associe une insuffisance rénale aigüe, un néphroblastome (tumeur de Wilms, 1/20 000 enfant) et, chez la plupart des individus XY concernés, une dysgénésie gonadique avec ambiguité génitale. Le syndrome de Frasier, lui, se caractérise par une néphropathie à évolution lente et une dysgénésie gonadique qui entraîne, chez les individus XY, un phénotype complètement féminin.
Le gène WT1 semble nécessaire à double dose (deux
allèles normaux) pour la détermination du testicule, alors
que le développement de l'ovaire peut se faire en présence
d'un seul allèle normal.
Chez la souris, l'invalidation du gène WT1 entraîne la
mort de l'embryon, qui, ne possédant ni rein ni gonades, présente
toujours un phénotype femelle par absence d'hormones mâles.
Ainsi le gène WT1 est impliqué dans la morphogénèse
de la crête urogénitale mais aussi dans la différenciation
testiculaire.
Comme WT1 et SRY sont tous les deux exprimés dans les cellules
de Sertoli, et qu'il existe, sur le promoteur du gène SRY, un site
capable de se lier avec une protéine WT1, on suppose que WT1
contrôle directement la transcription de SRY.
Enfin, l'une des isoformes protéiques du gène WT1 est
impliquée dans la transcription du gène de l'AMH.
Le gène SOX9
Il appartient à la famille des gènes SOX, puisqu'il code
pour une protéine (509 acides aminés) possédant un
domaine de fixation à l'ADN de type HMG présentant une forte
homologie (71% des aminoacides) avec celui de la protéine SRY. De
plus sa protéine possède (dans sa partie C-terminale) un
domaine impliqué dans l'activation de la transcription.
L'inactivation du gène SOX9 est responsable d'un syndrome: la
dysplasie campomélique. Les personnes atteintes présentent
des anomalies osseuses diffuses, et pour les trois-quarts des malades possédant
un caryotype 46/XY, des anomalies gonadiques. Ces dernières sont
diverses, allant de testicules à des ovaires dysgénésiques
avec quelques follicules primordiaux, ce qui constitue donc une inversion
sexuelle. Le phénotype externe est alors ambigu ou franchement féminin.
Le gène a été localisé sur le bras long
du chromosome 17 humain (17q).
Ce gène s'exprime dans les crêtes génitales des
souris des deux sexes. Ensuite, son expression augmente chez le mâle
dans les cellules de Sertoli au cours de la différenciation testiculaire
tandis qu'elle diminue dans l'ébauche ovarienne de la femelle, confirmant
ainsi son implication dans la différenciation gonadique mâle.
Le même dimorphisme de son expression existe chez d'autres
mammifères, des oiseaux, des reptiles, ce qui suggère pour
ce gène une fonction testiculaire majeure, conservée au cours
de l'évolution.
Des expériences in vivo sur des souris transgéniques ont
montré que la protéine SOX9 est indispensable à la
transcription de l'AMH. Dans les gonades foetales de mammifères,
l'expression de l'AMH se produit en effet après celle de SOX9. Cependant
dans d'autres taxons (poulet, certains reptiles), elle la précède.
Ainsi, la protéine SOX9 ne déclenche pas la lecture du gène
de l'AMH mais pourrait être responsable de l'élévation
de son expression dans le testicule foetal des vertébrés.
L'étude de patients de caryotype 46/XY, féminisés,
porteurs de mutations hétérozygotes de SOX9 ainsi que d'une
personne de caryotype 46/XX, masculinisée et porteuse d'une duplication
interstitielle du bras long du chromosome 17 a permis d'envisager
une intervention de la protéine SOX9 selon sa concentration. Un
taux élevé (trois allèles normaux, cas pathologique,
ou deux allèles, cas normal) de protéine SOX9 entraînant
une masculinisation, par activation de la transcription du gène
de l'AMH, alors qu'un taux faible (un seul allèle normal) n'assure
pas cette masculinisation.
Le gène DAX1
Dans l'espèce humaine, la duplication d'une région du chromosome X, localisée sur le bras court (Xp), entraîne le développement d'un phénotype féminin, avec dysgénésie gonadique, chez des individus de caryotype 46/XY. Cette région, de 160Kb, a été appelée DSS (Dosage Sensitive Sex-reversal) car elle contient au moins un gène capable, à double dose (deux allèles) d'inhiber la différenciation testiculaire. On a isolé un gène, DAX1, situé dans cette région et codant pour une protéine de 470 acides aminés présentant deux caractéristiques: sa partie C-terminale est similaire aux domaines de liaison du ligand des récepteurs nucléaires et sa partie N-terminale correspond à un nouveau type de domaine capable de se lier à l'ADN.
L'expression de ce gène présente, chez la souris, de grandes similitudes avec celles du gène sf1 ( en minuscules pour le gène murin) et il semble clair que ces deux facteurs soient impliqués dans le contrôle de la fonction de reproduction aux niveaux hypothalamo-hypophysaire et gonadique. Cette expression a été mise en évidence dans les crêtes génitales (et d'autres tissus: surrénales, hypothalamus et hypophyse) de souris des deux sexes, mais, si elle décroît ensuite dans les gonades mâles, elle se maintient chez les femelles, ce qui fait de ce gène un bon candidat à la fonction d'inversion sexuelle décelée dans la zone DSS.
Chez la souris, des lignées transgéniques surexprimant DAX1 (jusqu'à 5 fois la dose normale) ont été créées, et montrent effectivement un effet sur la différenciation testiculaire qui est alors retardée (mais pas empêchée). Chez l'homme des mutations du gène DAX1 sont responsables d'hypogonadotrophie.
Le gène SF1
Ce gène, localisé, chez l'homme, sur le bras long du chromosome
9 (9q), code pour une protéine appelée SF1, pour Stéroidogenic
Factor 1.
Cette protéine est un recepteur nucléaire d'un type particulier,
régulant la transcription d'un grand nombre de gènes d'enzymes
impliquées dans la stéroidogénèse des tissus
surrénalien et gonadique. SF1 régule également des
gènes d'hormones protéiques hypophysaire (sous-unité
bêta de la LH et la FSH) ou testiculaire (AMH en particulier).
Le facteur SF1 est trouvé très précocément
au cours du développement de la souris, chez des foetus de 9 jpc
(jours post-coïtum), au niveau des ébauches de gonades et ceci,
dans les deux sexes. Par la suite les profils d'expression diffèrent
selon les espèces.
Le gène de l'AMH
Il est localisé, dans l'espèce humaine, sur le chromosome
19. L'AMH (hormone anti-Müllérienne)
présente,
dans sa partie C-terminale qui doit être clivée au niveau
de la cellule cible pour devenir bio-active, une nette homologie avec certains
facteurs de croissance de la famille du TGF-bêta.
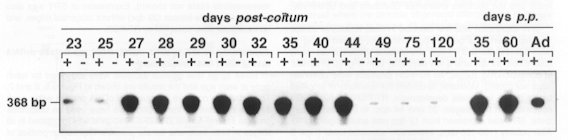
 |
|
chez le foetus de porc mâle (à gauche) et femelle (à droite) (Demande de droits en cours) Images obtenues par autoradiographie d'une RT-PCR d'ARN de gonades prélevées
à différents âges chez des foetus des deux sexes.
|
CONTROLE MULTIFACTORIEL DU GENE DE L'AMH
Vu l'influence de l'AMH dans la différenciation somatique, c'est le gène de cette hormone qui, par la régulation de son expression, va être responsable de l'engagement du tractus génital vers une voie mâle ou femelle.
Chez l'homme, le facteur SF1 régule la transcription de l'hormone anti-Müllérienne, mais SF1 n'agit pas seul, il fait partie d'un complexe de plusieurs facteurs de transcription, associant en particulier SOX9 et DAX1. De plus, l'une des isoformes protéiques du gène WT1 agit en synergie avec SF1 pour activer la transcription du gène de l'AMH.
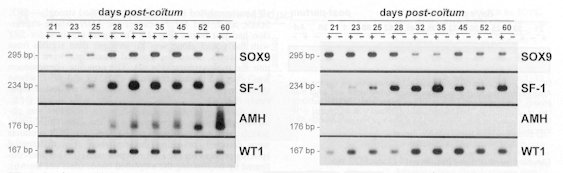
Il a été montré, in vivo, chez la souris, que des mutations dans les sites de fixation des protéines SOX9 et SF1, présents dans le promoteur du gène de l'AMH, entraînaient une modification de son expression. Chez cette espèce, Sox9 contrôle l'initiation de la transcription de ce gène et Sf1 le niveau de transcrits produits.
De plus, par des expériences de liaison protéique, on
a montré que les facteurs SOX9 et SF1 (humains) interagissent directement
l'un avec l'autre, SOX9 par son domaine de liaison à l'ADN, et SF1
par sa région terminale.
Par ailleurs, des expériences ont prouvé que la protéine
DAX1 est un inhibiteur de l'action de SF1 sur ce promoteur.
Ainsi, chez les femelles, un taux élevé de facteur DAX1
(ayant deux chromosomes X, elles possèdent ce gène en deux
exemplaires) inhiberait l'action de SF1 sur le promoteur du gène
de l'AMH. Ce gène ne serait donc pas transcrit.
A l'inverse, dans la gonade mâle, le taux élevé
de facteur SOX9 (gène autosomique) et faible de DAX1 (un seul chromosome
X) permettrait l'interaction du complexe de transcription SOX9/SF1 sur
le promoteur du gène de l'AMH, assurant ainsi, par la production
de cette hormone, une masculinisation des voies génitales.
BILAN DES CONNAISSANCES ACTUELLES
SUR LE DETERMINISME GENETIQUE DU
SEXE CHEZ LES MAMMIFERES
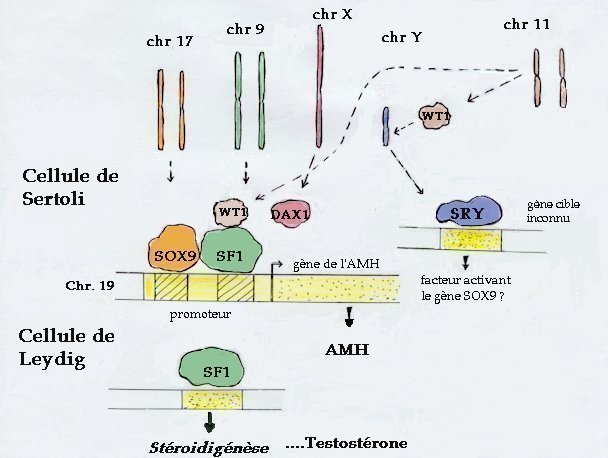 |
| Chez le mâle de mammifère, la détermination testiculaire
est déclenchée par SRY dans les précurseurs des cellules
de Sertoli. Le gène SOX9 est alors fortement transcrit et, en interaction
avec SF1, initie l'expression du gène de l'AMH. L'une des isoformes
protéiques du gène WT1 agit en synergie avec SF1 pour activer
la transcription du gène de l'AMH.
Dans les cellules de Leydig, le facteur SF1 active l'expression de gènes d'enzymes intervenant dans la stéroïdogénèse et de la testostérone est produite. Ces deux hormones testiculaires masculinisent alors les organes génitaux internes et externes. Chez la femelle, on ne connaît pas de gène déclenchant la différenciation ovarienne. On sait seulement que DAX1 inhibe l'action de SF1 et on a récemment identifié un gène (Wnt4) capable d'empêcher la différenciation des cellules de Leydig et leur production de testostérone (peut-être par l'intermédiaire de DAX1). D'autre part, chez la femelle, le niveau de transcription du gène SOX9 reste faible. |