Glossaire
Histoire
Téléchargements
Rédigée par Nadia Ouahioune, Lycée
Montesquieu, Le Plessis-Robinson
Relue par Paul Derer, Laboratoire Développement
et Vieillissement du système nerveux (J. Mariani)
Université Pierre et Marie Curie - Paris, UMR 7624 du CNRS
Cette synthèse a été rédigée à partir des ouvrages et articles suivants :
- Histologie moléculaire, J Poirier et
al., ed. Abrégés, MASSON, 1997 : 338-340
- Neuroanatomie, James D. Fix, En Bref
Ed. DeBoeck
Université, 1996, 105-112
- Derer Paul, Neuroscience letter, 1979 : 13
: 195-202
- Derer P, Derer M., Neuroscience 1990, 36 :
839-856
- Goffinet A. M., Médecine Science n°5,
vol 12, mai 1996
I - Organisation du cortex cérébral
Le cortex est une fine couche de substance grise qui recouvre les deux hémisphères et dont l'épaisseur varie entre 2 et 4 mm. Selon l'organisation histologique corticale, on distingue le néocortex qui représente 90% (constitué de 6 couches numérotées de I à VI de la surface vers la profondeur) et l'allocortex de structure plus simple (structure en 3 couches) qui ne représente que 10% de la surface total du cortex. Seul, le néocortex nous intéressera dans la mesure où il représente la plus grande partie du cortex.
1 - Les deux grandes classes de cellules corticales
Du point de vue cellulaire, on y distingue deux grandes classes : les cellules pyramidales (Cp) et les cellules étoilées (Ce) :
- les Cp ont une forme de cône, la pointe étant dirigée vers la surface méningée. Le dendrite principal de ces cellules se dirige vers la surface tandis que l'axone se dirige vers la substance blanche.
- Les Ce ont un corps cellulaire plus petit. L'arborisation dendritique s'étend sur toute la surface du corps cellulaire donnant ainsi un aspect étoilé.
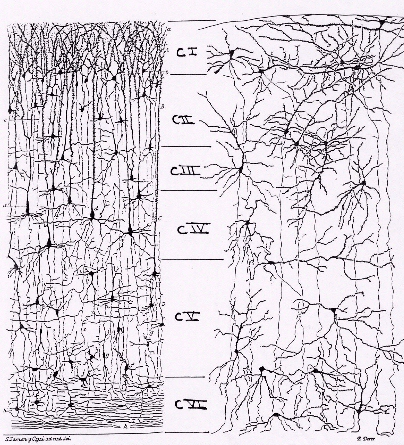
2 - Les six couches du néocortex constituant la substance grise
Horizontalement, on distingue 6 couches numérotées
de I à VI de la surface vers la profondeur.
|
Le cortex est formé d'une majorité de neurones pyramidaux:
|
Le cortex est formé d'une majorité de de neurones pyramidaux :
|
 |
|
Horizontalement, on distingue 6 couches numérotées de I à VI de la surface vers la profondeur :
- la couche I ou couche moléculaire pauvre en cellules nerveuses, constituée surtout des cellules gliales et des fibres nerveuses à trajets parallèles à la surface corticale.
- la couche II ou couche granulaire externe, essentiellement constituée de petits neurones non pyramidaux.
Remarque : chez les rongeurs, ces deux couches (I et II) n'en forment qu'une.
- La couche III ou couche pyramidale externe, contient la majorité des neurones pyramidaux. Les axones de ces neurones forment les fibres intrahémisphériques et interhémisphériques ou commissurales dans la substance blanche tandis que les dendrites apicaux atteignent la couche moléculaire.
- La couche IV ou couche granulaire interne formée de petits neurones non pyramidaux représentés essentiellement par des cellules étoilées.
- La couche V ou couche pyramidale interne contient essentiellement des neurones pyramidaux dont les dendrites apicaux se projettent soit dans la couche moléculaire (couche I) soit dans la couche granulaire interne (couche IV). Les axones de ces neurones se projettent à distance (striatum, tronc cérébral, moelle épinière..) contrairement à ceux de la couche III.
- La couche VI ou multiforme est la source principale des neurones qui se projettent sur le thalamus. Elle est à l'origine aussi des fibres commissurales et des fibres d'association.
II - Organisation du cortex cérébelleux
A l'inverse du cortex cérébral, le cortex cérébelleux n'est constitué que de trois couches : une couche externe ou moléculaire, la couche des Cellules de Purkinje réparties en seule assise et la couche interne ou granulaire. L'organisation du cortex cérébelleux a été largement développée dans "Structure du cervelet chez la souris sauvage".
III- Développement du cortex cérébral
1 - Chez la souris normale
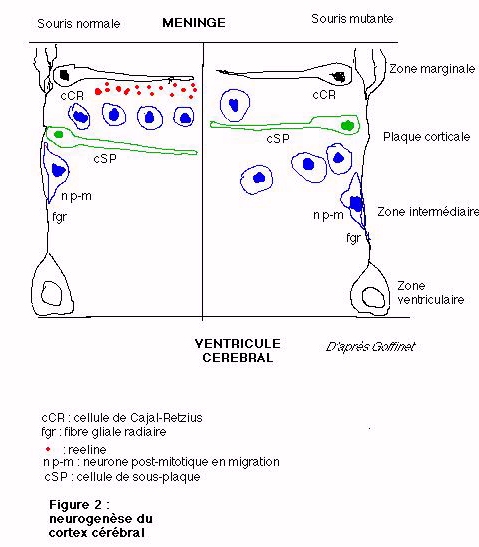
Dès le stade ftale, une préplaque corticale composée des premiers neurones postmitotiques engendrée au sein des zones ventriculaires, se met en place. Ces cellules migrent ensuite le long de fibres gliales radiaires, jusqu'à la surface méningée et deviennent soit des cellules de Cajal-Retzius (cCR) soit des cellules de la sous-plaque (SP). La plupart de ces cellules disparaîtront lorsque les neurones de la plaque corticale se mettront en place entre la couche des cCR (désormais située dans la zone marginale) et de la SP. Enfin la plaque corticale se transformera en substance grise du cortex définitif (décrite au dessus).
La neurogenèse de la plaque corticale se réalise donc selon un gradient intéro-externe de migration amenant les derniers neurones formés dans les zones ventriculaires à occuper toujours au terme de leur migration, l'interface entre la couche marginale et la plaque corticale. Le développement de cette plaque est très important car il est à l'origine des six couches du cortex mature; la zone ventriculaire ne constituant plus à la fin des processus de corticogenèse qu'une couche unique de cellules épendymaires qui limite les ventricules cérébraux.
La reeline, protéine extracellulaire sécrétée par les cCR est déterminante dans la mise en place des couches de neurones juxtaposés selon ce gradient intéro-externe. Ce rôle dans la corticogenèse pourrait être une action répulsive, empêchant les neurones en migration de pénétrer dans la zone marginale riche en reeline respectant ainsi la stratification normale du cortex mature (figure 2). Malgré le rôle essentiel de cette protéine dans la corticogenèse, elle n'est qu'un maillon important de la cascade des interactions moléculaires présidant à la mise en place des couches corticales.
2 - Chez la souris homozygote reeler
Chez l'embryon de la souris homozygote reeler, la migration des neurones se déroule normalement jusqu'au moment où ceux-ci arrivent près de leur destination. La reeline n'étant pas sécrétée dans la matrice extracellulaire par les cellules de Cajal-Retzius, le gradient répulsif vis-à-vis des neurones de la plaque corticale n'a pas lieu si bien que les cellules de la sous-plaque sont comme repoussées vers l'extérieur (figure 2) en dehors de la plaque corticale sous-jacent très désorganisée. Cela se traduit chez l'animal très précocement par un cortex où les couches sont peu apparentes (figure 1b) : la plaque corticale ne s'intercale plus dans la préplaque et le gradient de mise en place des neurones pyramidaux se fait de manière quasiment inversée c'est-à-dire selon un gradient extéro-interne.
Remarque : dans des zones cérébrales où la reeline est très exprimée chez la souris normale, son absence chez le mutant reeler n'entraîne pas de perturbation importante, c'est notamment le cas pour le bulbe olfactif mais pas pour le cervelet (voir ci-dessous).
IV - Développement du cortex cérébelleux
1 - Chez la souris normale
Dans le cortex cérébelleux, le rôle joué par les cellules de Cajal-Retzius dans le cortex cérébral en formation, serait joué par les cellules granulaires externes. En effet, ces cellules sécréteraient la reeline dans la matrice extracellulaire qui aurait un rôle répulsif dans la zone marginale repoussant la plaque des cellules de Purkinje à l'interface de la couche moléculaire et de la couche granulaire.
2 - Chez le mutant homozygote reeler
Dans le cortex cérébelleux où la reeline est pourtant peu exprimée, la modification de la séquence nucléotidique de cette protéine entraîne de grandes perturbation dans l'organisation du cortex cérébelleux chez le mutant homozygote. Chez ce mutant, les cellules de Purkinje sont disposées de manière aléatoire vraisemblablement à cause d'un arrêt de leur migration si bien que le cervelet paraît constitué de l'emboîtement de deux structures : à l'extérieur, un cortex cérébelleux dont l'architecture est semblable à celle que l'on trouve chez l'animal normal mais très mince et à l'intérieur une masse cellulaire comportant la plupart des cellules de Purkinje mélangées aux cellules des noyaux profonds. Faire un renvoi vers les photos et la synthèse correspondant à cette partie.
3 - Chez d'autres mutants cérébelleux
Les précurseurs des cellules granulaires du cortex cérébelleux, localisés dans la couche moléculaire superficielle, mettent d'abord en place leurs axones (futures fibres parallèles) avant que leurs corps cellulaires ne migre vers leur position définitive dans la couche granulaire où elles contacteront les fibres moussues.
Chez la souris mutante weaver (qui comme le reeler et le staggerer présente des modifications du phénotype à l'échelle clinique et moléculaire), les cellules précurseurs des grains meurent durant les deux premières semaines postnatales avant qu'elles n'aient pu migrer dans la couche granulaire interne et former leurs fibres parallèles. Secondairement à la mort cellulaire massive des cellules précurseurs des grains, les fibres moussues ne trouvant plus leur cible forment alors des synapses avec les cellules de Purkinje. Cette mutation est donc à l'origine de la désorganisation des circuits neuroniques par disparition de leur cible.
Il en est de même pour le mutant homozygote staggerer chez qui les cellules précurseurs des cellules granulaires migrent vers la couche moléculaire mais disparaissent presque totalement un mois après la naissance à cause de la mort de leur cible, les cellules de Purkinje.
Outre l'intérêt des mutants cérébelleux dans la diversité des phénotypes, ces trois mutants (staggerer, weaver et reeler) sont à l'état homozygote de bons modèles pour étudier l'implication du génotype dans la mise en place des réseaux neuroniques et par voie de conséquence du fonctionnement du système nerveux.