
Eploitation pédagogique - ASPM
Le gène ASPM et la spécificité du cerveau humain
1 - Un gène fréquemment cité à propos de l’évolution du cerveau
La démarche classique consistant à comparer le développement de l'Homme et du Chimpanzé met l’accent sur la période prolongée de multiplication des neurones chez l’Homme par rapport au Chimpanzé. On a ainsi une explication au niveau cellulaire de l’importance du volume du cerveau humain et particulièrement des hémisphères cérébraux (et donc du cortex) par rapport au Chimpanzé. Ensuite, on passe au déterminisme génétique de cette différence et il est habituel de faire intervenir le gène ASPM. Ainsi, dans l’accompagnement en ligne d’un manuel de terminale, on peut lire : « La protéine codée par le gène ASPM détermine la durée de la phase de multiplication des cellules souches du cortex cérébral. Différentes mutations de ce gène se caractérisent donc par un cortex cérébral plus ou moins important. Un tel gène a donc pu jouer un rôle important dans l’histoire évolutive de notre espèce. »
Nous allons essayer de préciser un peu les affirmations de cet extrait, en examinant notamment les arguments qui corroborent l’hypothèse de l’implication de ce gène dans l’accroissement du cerveau dans la lignée humaine. La méthodologie qui a été suivie est celle des gènes candidats (voir Génétique moléculaire et spécificités humaines - la méthode des gènes candidats), celle déjà utilisée pour FOXP2.
2 - Un gène à l’origine d’anomalies cérébrales dans l’espèce humaine
Une anomalie d’origine génétique est la microcéphalie caractérisée par une réduction de 2 à 4 fois de la circonférence de la tête, associée à une réduction correspondante du volume cérébral, en particulier des hémisphères cérébraux. L’organisation cérébrale est par ailleurs quasi normale. Cette anomalie s’accompagne d’un retard mental léger à modéré. Les autres organes du corps ne sont pas affectés : c’est donc une anomalie spécifique du cerveau.
L’analyse génétique dans les familles ayant des membres atteints de ce syndrome a montré qu’il s’agissait d’une maladie génétique monogénique, autosomale récessive. Les chercheurs ont découvert que plusieurs loci peuvent être à l’origine de ce syndrome (7 ont été identifiés) dont le gène ASPM situé sur le chromosome 1. MCPH est le symbole général donné à ces gènes ; ASPM est ainsi le gène MCPH5. C’est celui qui est le plus fréquemment impliqué dans la microcéphalie.
On connaît de nombreuses mutations du gène ASPM ; beaucoup d’entre elles étant des mutations non-sens entraînant l’apparition anticipée d’un codon stop et donc la formation d’une protéine tronquée, non fonctionnelle. Voici le pedigree d'une famille avec des individus atteints de microcéphalie.
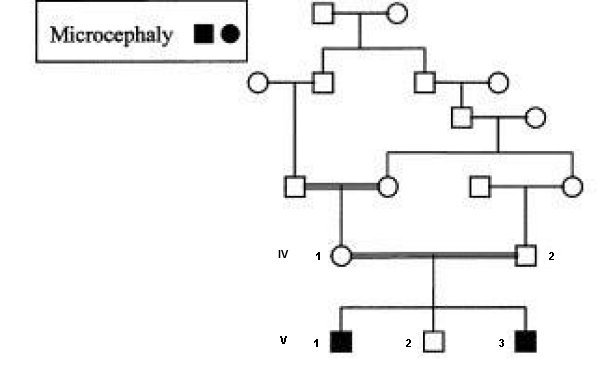
La microcéphalie est une maladie héréditaire très rare dont la prévalence est plus grande dans les populations asiatiques et arabes où des mariages consanguins ont lieu. Dans une population du Yorkshire (GB) l'incidence est de 1 par million dans la population européenne et de 1 sur 10 000 dans une population originaire du nord du Pakistan. La famille représentée est d'origine pakistanaise.
3 - Le rôle de la protéine ASPM
ASPM signifie : Abnormal Spindle-like Microcephaly-associated ; cela indique que la protéine intervient dans la formation et la structure du fuseau de division au cours des mitoses. Les études chez la Souris ont montré qu’il s’exprime fortement durant la période de neurogénèse cérébrale. C’est son seul territoire d’expression. On peut donc comprendre qu’il agisse sur les divisions cellulaires créatrices de la population de neurones du cerveau, mais les modalités précises de son action sont inconnues. En tout cas, il n’est pas prouvé qu’il détermine la durée de la phase de multiplication des cellules souches des neurones.
4 - L’hypothèse sur l’importance évolutive du gène ASPM
Les mutations du gène ASPM qui rendent non fonctionnelle la protéine ASPM chez les humains actuels ont pour conséquence la réduction du volume cérébral à une valeur proche de celui des australopithèques. Les chercheurs ont alors fait l’hypothèse que dans la lignée humaine, il y avait eu une évolution particulière du gène ASPM ayant pu contribuer à l’accroissement du volume cérébral.
Un test de cette hypothèse
Pour tester cette hypothèse diverses équipes de chercheurs ont séquencé le gène ASPM chez divers primates puis ont comparé les séquences de ces gènes homologues. On peut le faire avec Anagène et comparer non seulement les séquences codantes du gène mais aussi les protéines. C’est la même démarche qu’avec FOXP2. Cependant l’interprétation des données est plus délicate car c’est un gène très long (10434 bp pour la partie codante et 3477 acides aminés pour la protéine) et il y a eu de nombreuses mutations dans les diverses lignées. Cela ne rend pas immédiatement apparente l’originalité de l’évolution du gène dans la lignée humaine.
Pour exploiter les données, il faut dénombrer le nombre de mutations survenues dans les différentes branches de l’arbre phylogénétique des primates. Plus précisément, il faut dénombrer le nombre de substitutions silencieuses et de substitutions faux sens (conduisant à un changement d’acide aminé) survenues dans chaque lignée. Vu la longueur du gène, c’est un travail long et assez difficile. On peut se contenter avec Anagène de comparer simultanément les séquences nucléiques et protéiques chez l’Homme, le Chimpanzé et le Gorille. Cela suffit pour dénombrer les mutations survenues dans la lignée humaine et dans celle du Chimpanzé mais non dans celle du Gorille.
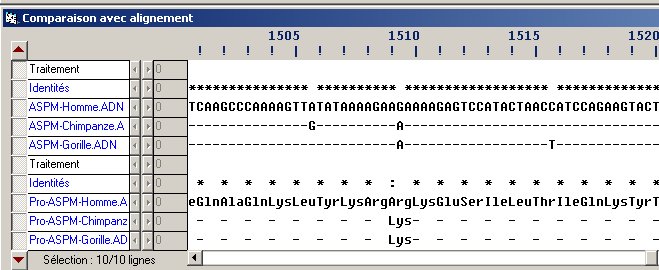
Portion de l'alignement des séquences nucléiques et des séquences protéiques correspondantes du gène ASPM chez l'Homme, le Chimpanzé et le Gorille. Illustration d'une mutation silencieuse et d'une mutation non silencieuse. La première mutation silencieuse à gauche a eu lieu chez le Chimpanzé puisque l'Homme et le Gorille ont le même nucléotide à ce site. La mutation faux sens a eu lieu dans la lignée humaine puisque à ce site l'Homme possède un nucléotide différent de ceux du Chimpanzé et du Gorille (même nucléotide chez ces deux taxons). A noter que l'on ne peut affirmer sur cette seule comparaison que la mutation chez le Gorille a été acquise dans la lignée du Gorille, en l'absence d'informations sur ce site chez l'Orangutan, le Gibbon, le Macaque, etc.
On peut comparer le nombre de mutations silencieuses et faux sens dans les deux lignées (celle de l'Homme et du Chimpanzé) et calculer le rapport mutations faux sens/mutations silencieuses pour chacune d'elles.
|
Substitutions silencieuses |
Substitutions faux-sens |
Total |
|
|
Lignée humaine |
7 |
19 |
26 |
|
Lignée du Chimpanzé |
10 |
15 |
25 |
Ce rapport est plus élevé dans la lignée humaine (2,7) que dans celle du Chimpanzé (1,5). Cela indique que dans la lignée humaine, les mutations entraînant un changement d’acide aminé se sont davantage fixées que dans la lignée du Chimpanzé. Cela suggère qu’elles ont été sélectionnées car elles devaient conférer aux organismes qui en étaient porteurs un avantage sélectif
Remarque :
Le raisonnement précédent manque de rigueur car ce qu’il faut calculer c’est le rapport entre taux de substitutions faux-sens (non synonymes) sur le taux de substitutions silencieuses (synonymes). En effet dans un gène, du fait des caractéristiques du code génétique, il y a un nombre maximal de sites dont la mutation peut être silencieuse. En première approximation le nombre de sites potentiellement silencieux est entre 20% et 35% du nombre de sites total du gène. Et pour les mutations non synonymes le nombre maximal de sites est compris entre 65% et 80% approximativement. Il faut alors calculer les rapports Ka et Ks.
Ka : nombre de substitutions non synonymes observé / nombre maximal de substitutions non synonymes possible.
Ks : nombre de substitutions synonymes observées / nombre maximal de substitutions synonymes possibles.
Il reste alors à calculer le rapport Ka/ks
Si Ka/ks est égal à 1 cela signifie que les substitutions non synonymes se sont fixées à la même vitesse que les substitutions synonymes. Comme celles-ci sont neutres, cela signifie que les mutations non synonymes l’étaient aussi et n’ont pas conféré un avantage sélectif. Elles ont été fixées sans doute par dérive génique.
Si le rapport ka/ks est nettement inférieur à 1, cela signifie que les substitutions non synonymes se sont fixées moins vite que les substitutions synonymes, ce qui indique que la plupart d’entre elles ont été sélectionnées négativement (elles devaient être plus ou moins délétères pour les organismes qui les possédaient).
Si le rapport Ka/ks est supérieur à 1 cela indique que les substitutions non synonymes se sont davantage fixées que les substitutions synonymes. C’est le signe d’une sélection positive : elles devaient conférer aux organismes porteurs un avantage sélectif par rapport à ceux qui ne les possédaient pas.
5 - Bilan
L’arbre phylogénétique des primates ci-dessous indique le rapport Ka/Ks pour le gène ASPM sur les diverses branches. D'après Evans.

On constate un rapport supérieur à 1 sur la branche conduisant aux hominidés et surtout sur celle de la lignée humaine. Les substitutions non synonymes de ce gène ont donc été sélectionnées positivement, particulièrement dans la lignée humaine. Cela corrobore l’hypothèse que les mutations dans la lignée humaine ont pu contribuer à l’accroissement du cerveau. On peut penser que l’accroissement du volume cérébral qu’elles pouvaient provoquer dotait les organismes de nouvelles aptitudes leur conférant un avantage sélectif. Mais on n’a jusqu’à présent nullement montré en quoi ces mutations changeaient les propriétés de la protéine ASPM. Le seul indice du rôle des mutations du gène ASPM dans l’accroissement du volume cérébral dans la lignée humaine est donc la détection d’une sélection positive.
A titre de conclusion les deux arbres ci-dessous visualisent les moments de sélection positive dans l’histoire évolutive des gènes FOXP2 et ASPM chez les Primates.
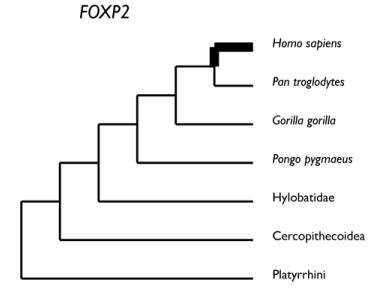
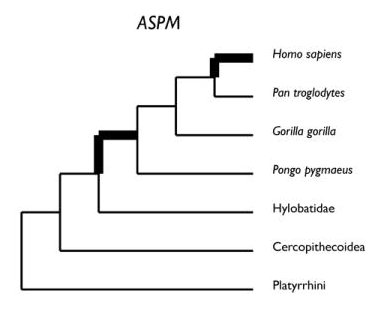
D'après Vallender. En gras les périodes pendant lesquelles on a mis en évidence l'existence d'une sélection positive entraînant un changement dans la séquence de la protéine codée par le gène. On suppose que ces changements ont pu jouer un rôle dans la genèse des circuits neuroniques du langage pour FoxP2, et de l'accroissement du cerveau pour ASPM.