
Génétique de l’adaptation à l’altitude des Tibétains
Génétique de l’adaptation à l’altitude des Tibétains
L’implication d’un gène : EPAS1
Durant la première décennie du XXIème siècle, les chercheurs ont bénéficié d’une amélioration considérable des techniques de séquençage de génomes entiers et de celles permettant l’exploitation des séquences obtenues. Grâce à cela, une équipe de chercheurs publie en 2010 les résultats d’une étude sur la génétique de l’adaptation à l’altitude des Tibétains.
Ces chercheurs ont séquencé le génome entier de 50 Tibétains vivant à une altitude de 4300 mètres, et de 40 Chinois Hans de Beijing (Pékin) dont l’altitude est inférieure à 50 mètres. Ces deux populations sont étroitement apparentées et la base du raisonnement tenu par les chercheurs est le suivant : si on trouve des différences importantes à certains endroits du génome de ces deux groupes, on pourra supposer que ces régions ont une relation avec l’adaptation à l’altitude ; plus exactement, que les caractéristiques génomiques propres aux Tibétains leur assurent de pouvoir vivre à des altitudes élevées sans souffrir du mal des montagnes.
La variabilité prise en compte par les chercheurs est celle des SNP (Single Nucléotide Polymorphism). Ce sont des sites de la séquence de l’ADN où il existe au moins deux variants dans la population (par exemple un nucléotide à adénine et un nucléotide à guanine). Par extension, on parle d’ « allèle » propre à chaque nucléotide d’un SNP. Ces SNP, au nombre de 10 millions environ dans le génome humain, peuvent être situés dans les régions codantes des gènes (exons), dans les introns et dans les régions inter-géniques. Les chercheurs ont évalué pour chaque SNP, la fréquence de chaque « allèle » dans chacune des deux populations. Le graphique ci-dessous traduit les données obtenues.
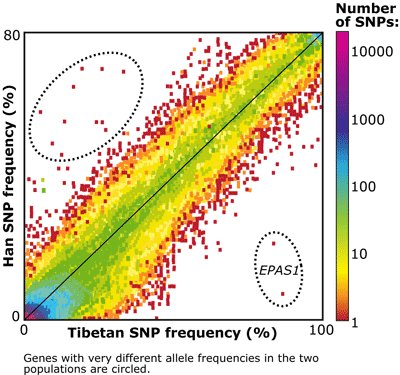
D'après Yi X, Liang Y, Huerta-Sanchez E, et all. Sequencing of 50 human exomes reveals adaptation to high altitude. (Modifié).
Sur ce graphique, chaque point représente un SNP ou plus exactement un « allèle » de ce SNP. En abscisse se trouve la fréquence de cet allèle chez les Tibétains (de 0 à 100%) et en ordonnée celle chez les Hans (de 0 à 80%). L'échelle montre que les points rouges représentent des allèles unitaires (1 seul allèle d'un SNP). Les SNP dans les régions violettes, bleues, vertes, jaunes ne sont pas individuellement identifiables vu leur nombre. La couleur violette par exemple indique qu'un très grand nombre d'allèles de SNP ont une très faible fréquence dans les deux populations.
Le fait le plus frappant de ce graphique est qu’on constate que la fréquence de la majorité des allèles des SNP des Tibétains et des Hans est la même : le nuage de points autour de la diagonale du graphe le traduit. Cela confirme l’étroite parenté des deux populations. Toutefois, certains SNP s’écartent de cette covariance. Certains allèles de SNP ont une plus forte fréquence chez les Hans que chez les Tibétains. Vu leur faible fréquence chez ces derniers, on peut penser qu’ils ne jouent aucun rôle dans l’adaptation à l’altitude de cette population. En revanche, on constate dans la région du génome correspondant au gène EPAS1 l'existence de deux SNP pour lesquels « un allèle » a une fréquence nettement plus élevée chez les Tibétains que chez les Hans. Pour le SNP où la différence entre les deux populations est la plus forte, la fréquence de l’allèle G est de 87% (et donc celle de l’autre allèle C de 13%) contre 9% chez les Hans pour l'allèle G (91% pour l’allèle C). Ce SNP discrimine nettement les deux populations et on peut supposer qu’il est impliqué dans l’adaptation à l’altitude des Tibétains.
Encore faut-il qu’il soit un allèle dérivé. Les chercheurs ont analysé le génome d’une population de Danois et ont constaté qu’à ce SNP, la fréquence de l’allèle C est de 100%. Autrement dit les Danois possèdent uniquement l’allèle très majoritaire chez les Hans. Cela indique que cet allèle est l’allèle ancestral et l’allèle G des Tibétains l’allèle dérivé. Dans une perspective évolutive, cela signifie que l’allèle G s’est répandu au cours de l’histoire des populations de Tibétains, sans doute sous l’action de la sélection naturelle, car il conférait un avantage sélectif aux personnes qui le possédaient.
Pour tester l’idée que le gène EPAS1 est bien en jeu dans l’adaptation à l’altitude des Tibétains, les chercheurs ont déterminé le nombre d’hématies et la concentration d’hémoglobine chez les Tibétains en fonction de leur génotype.
| Génotype | Nombre de Tibétains présentant le génotype | Concentration moyenne en Hémoglobine (g/l) | Nombre moyen d'hématies |
| CC | 10 | 178 | 5,3 |
| CG | 84 | 178,9 | 5,6 |
| GG | 272 | 167,5 | 5,2 |
On constate que les Tibétains homozygotes pour « l’allèle » dérivé G ont le nombre d’hématies et la concentration d’hémoglobine les plus faibles, traits caractéristiques de l’adaptation à l’altitude des Tibétains. Bien que les différences avec les deux autres génotypes ne soient pas très fortes, les auteurs estiment que cela confirme le rôle joué par EPAS1 chez les Tibétains.
Il faut bien remarquer que le polymorphisme du SNP caractéristique des Tibétains se trouve dans un intron et que les auteurs n’ont pas déterminé comment il pouvait agir. Ils suggèrent une répercussion sur l’épissage du gène.
EPAS1 des Tibétains et les denisoviens
La même équipe de chercheurs souhaitant faire une analyse plus poussée, a séquencé de nouveau la région de l’ADN contenant la séquence entière de EPAS1 (région régulatrice en amont, exons et introns) chez deux populations, l’une de Tibétains, l’autre de Chinois Hans. Ils ont identifié dans un intron, une région de 2500 paires de base remarquable par 5 sites, 5 SNP polymorphes. L’association d’allèles de SNP situés sur la même portion de la molécule d’ADN forme un haplotype.
On dispose des séquences de cette région de 2500 Kb trouvées chez les Tibétains et les Hans (Fichier TIB1-TIB2-Han.edi). A partir de la comparaison de ces séquences, on peut déterminer les haplotypes dans les deux populations. Chaque haplotype est défini par l’association d’allèles des 5 sites.
Le graphique ci-dessous indique la fréquence de ces haplotypes dans les deux populations.

D'après : Altitude adaptation in Tibetans caused by introgression of Denisovan-like DNA. Nature 512, 194–197 (14 August 2014)
On constate une différence entre les deux populations : chez les Tibétains la présence de deux haplotypes, l’un AGGAA, l’autre GAAGG ; chez les Hans on n’a trouvé qu’un seul haplotype GAAGG. Surtout, chez les Tibétains, la fréquence de l’haplotype non trouvé chez les Hans (AGGAA) est de plus de 80%. Cela confirme une différence très forte entre les deux populations au niveau du gène EPAS1 et corrobore l’implication de ce gène dans l’adaptation à l’altitude des Tibétains.
Les chercheurs ont ensuite identifié les haplotypes relatifs à cette séquence EPAS1 de 2,5 kb présents dans différentes populations mondiales, grâce aux données du 1000 genomes project (Haplotypes Epas1.edi). Le graphique ce-dessous renseigne sur les résultats obtenus.
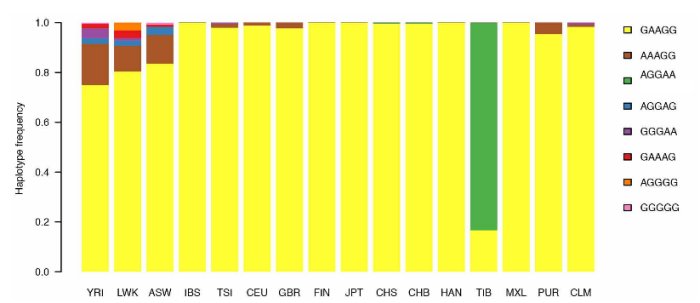
D'après : Altitude adaptation in Tibetans caused by introgression of Denisovan-like DNA. Nature 512, 194–197 (14 August 2014)
Nomenclature des populations concernées (1000 genomes project)
En dehors de deux Chinois Hans, l’un de Chine du sud (CHS), l’autre de Beijing( CHB), on constate que l’haplotype très largement représenté chez les Tibétains est absent de toutes les populations mondiales. Il est spécifique aux Tibétains et ainsi corrélé à leurs caractéristiques phénotypiques.
Les auteurs ont recherché une explication à cet haplotype propre aux Tibétains. On peut constater qu’aux 5 sites, les nucléotides de cet haplotype (AGGAA) sont différents de ceux de l’haplotype très majoritaire dans les autres populations (GAAGG) ; cet haplotype est donc ancestral.
Dans l’interprétation classique « mutation - sélection », cela suppose que 5 mutations indépendantes soient survenues sur la même molécule d’ADN et que l’haplotype résultant se soit ensuite généralisé à la population de Tibétains sous l’action de la sélection naturelle. La comparaison avec les génomes d'autres Homo a suggéré une autre explication.
Depuis 2010, on dispose d’une version de la séquence nucléaire de l’homme de Neandertal, version qui a été ensuite précisée dans une publication en 2014. En outre, en 2012, une phalange a été trouvée dans la grotte de Denisova en Sibérie, dont on a réussi à extraire l’ADN nucléaire et à le séquencer. Cet ADN nucléaire est manifestement celui d’un Homo mais différent des sapiens et des néandertaliens. Bien qu’on ne connaisse rien de son squelette, on a créé sur la base des caractéristiques de cet ADN fossile, un autre groupe d’Homo, celui des denisoviens, groupe aujourd’hui éteint, comme celui des néandertaliens.
On dispose des séquences homologues des haplotypes des sapiens chez ces Homo fossiles. On peut alors les comparer avec celles des Tibétains (Denisovien-Néandertalien-Tibétains.edi). On s’aperçoit que l’haplotype trouvé chez les denisoviens qui ont été séquencés est identique à celui fréquent chez les Tibétains et absent des autres populations de sapiens. En revanche, l'haplotype spécifique des Tibétains (AGGAA) ne se retrouve pas chez les néandertaliens (GGGAG). Cela conduit à une autre explication sur l’origine de l’haplotype des Tibétains. On imagine, il y a plus de 30000 ans des rencontres en Asie entre des denisoviens et des sapiens, rencontres suivies d’hybridations ayant conduit à l’intégration de fragments d’ADN de denisoviens chez cette population de sapiens. Les denisoviens devaient posséder les caractéristiques génétiques du gène EPAS1 adaptant l’organisme à la haute altitude. Cette introgression a dû avoir lieu avant la séparation de ce groupe humain en deux groupes, celui des Hans et celui des Tibétains, il y a 20000 ans environ. L’allèle EPAS1 (au sens allèle d’un gène) devait être peu fréquent au début dans la population de Tibétains mais, lorsque ceux-ci se sont établis à de hautes altitudes, il s’est répandu au fil des générations sous l’action de la sélection naturelle. Par contre dans la population des Hans restés à basse altitude, la possession de cet allèle n’apportait aucun avantage sélectif et il y a quasiment disparu.