
Famille multigénique des hormones hypophysaires et placentaires
Gènes des hormones hypophysaires et placentaires
Informations scientifiques
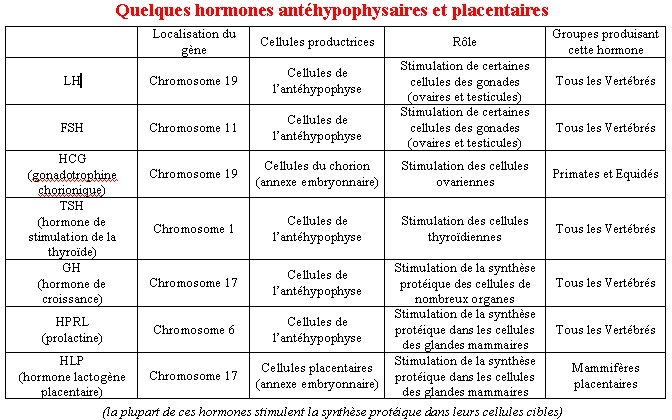
Les hormones antéhypophysaires et placentaires forment deux familles multigéniques : la famille des gènes LH, FSH, TSH et HCG et celle des gènes GH, HPRL et HLP.
La famille des gènes LH, FSH, TSH et HCG
Les hormones antéhypophysaires LH, FSH et TSH et l’hormone HCG sont des glycoprotéines formées par l’assemblage de deux chaînes polypeptidiques, les chaînes alpha et bêta. Ces quatre hormones diffèrent uniquement par leur chaîne bêta qui est la chaîne responsable de la spécificité fonctionnelle de ces hormones.
Ces hormones jouent des rôles différents dans l’organisme : les hormones LH et FSH participent au contrôle hormonal du fonctionnement de l’appareil reproducteur chez l’homme et chez la femme, l’hormone TSH stimule le fonctionnement thyroïdien et l’hormone HCG stimule le corps jaune lors d’un début de grossesse.
Ces chaînes bêta sont codées par des gènes qui sont situés sur des chromosomes différents (chromosome 1 pour le gène de TSH bêta, chromosome 11 pour le gène de FSH bêta et chromosome 19 pour les gènes de LH bêta et de HCG bêta).
Plusieurs arguments permettent de dire que ces gènes forment une famille multigénique :
-
ces quatre gènes présentent la même structure globale (trois exons et deux introns) ;
-
ces quatre gènes et les protéines pour lesquelles ils codent présentent des similitudes de séquences assez importantes ;
-
au niveau protéique, on peut constater la présence de douze résidus cystéine à des sites identiques pour les quatre hormones. Or, ces résidus, en permettant la formation des ponts disulfures, jouent un rôle essentiel dans la stabilisation de la structure tridimensionnelle de ces chaînes et donc de leurs propriétés.
On peut donc considérer que les similitudes constatées au niveau protéique et nucléique traduisent une origine commune pour toutes ces hormones et les gènes qui les codent. Ces gènes sont issus d’un même gène ancestral, et ils sont apparus par duplications géniques suivies de mutations indépendantes. La duplication la plus récente est celle qui est à l’origine des gènes LH bêta et HCG bêta.
Ce qu’il est important de noter ici, c’est que la divergence des copies dupliquées a conduit à des gènes codant pour des polypeptides ayant des fonctions différentes.
Tous les Vertébrés possèdent le gène de LH bêta, mais seuls les Mammifères placentaires (notamment les Primates et les Équidés) possèdent le gène de HCG bêta. On peut donc penser que le gène HCG bêta provient d’un gène LH bêta ancestral, la duplication étant postérieure à l’apparition des Mammifères placentaires.
style="margin-left:0.35pt; text-align:justify">La comparaison des protéines LH bêta et HCG bêta montre que ces deux protéines n’ont pas la même longueur : 165 acides aminés pour HCG bêta et 141 acides aminés pour LH bêta (méthionine comprise). Or, la comparaison des séquences nucléiques des gènes de ces deux hormones montre une délétion d’un nucléotide (en position 402) ; cette délétion entraîne un décalage du cadre de lecture lors de la traduction, décalage qui fait disparaître le codon stop présent dans le gène de LH (codon n° 142). C’est cette mutation par délétion qui est donc responsable de l’allongement de la protéine HCG bêta par rapport à la protéine LH bêta.
Remarque
La duplication du gène ne concerne pas uniquement la séquence strictement codante mais aussi celle qui se trouve en amont et en aval de celle-ci. Pour faciliter la compréhension de l’allongement de la séquence codante de HCG (et donc de sa protéine) par rapport à LH, nous avons fourni une séquence appelée LHADNC constituée par la séquence codante de LH, suivie d’une séquence de nucléotides homologue de la fin de celle de HCG mais non transcrite dans LH. La comparaison de LHADNC et de HCG permet d’expliquer que la protéine HCG a 24 acides aminés de plus que LH : par suite de la délétion, le gène HCG incorpore dans sa séquence codante la région en aval de LH non transcrite.
On peut conduire les élèves à la représentation suivante :
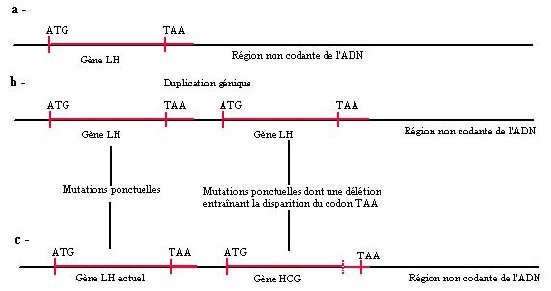
a - Situation initiale il y a plus de 60 Ma dans une espèce ancestrale des Primates.
b – Duplication génique de LH bêta.
c - Situation actuelle : la délétion dans le duplicata, en entraînant un décalage du cadre de lecture, a fait disparaître le codon stop. La région codante du gène incorpore une partie de l’ADN non codant situé en aval, jusqu’à un nouveau codon stop.
En toute rigueur, pour bien comprendre tout cela, il faut savoir que l’ARNm comporte une région normalement non codante après le codon stop UAA. Si ce codon stop disparaît, l’ARNm est traduit jusqu’à la rencontre d’un nouveau codon stop.
La famille des gènes GH, HPRL et HLP
Les hormones GH (hormone de croissance produite par l’antéhypophyse), HPRL (hormone prolactine, produite par l’antéhypophyse) et HLP (hormone lactogène placentaire produite par le placenta) ont des actions différentes mais apparentées. La HLP agit sur la glande mammaire en stimulant la synthèse protéique, comme la HPRL (toutefois plus efficace), et la GH stimule également la synthèse protéique dans de nombreux tissus.
Ces hormones sont codées par des gènes situés sur des chromosomes différents : chromosome 6 pour le gène de la HLP, chromosome 17 pour les gènes de la GH et la HPRL.
Les similitudes de séquences nucléiques et protéiques constatées permettent de dire que ces gènes forment une famille multigénique :
Ces trois gènes ont donc une origine commune et sont issus d’un même gène ancestral par duplications géniques suivies de mutations différentes.
La duplication la plus récente est celle qui est à l’origine des gènes GH et HLP.
Tous les Vertébrés tétrapodes possèdent les gènes GH et HPRL, mais seuls les Mammifères placentaires possèdent le gène HLP.
On peut donc considérer qu’une première duplication (à l’origine des gènes GH et HPRL) a eu lieu chez les premiers tétrapodes, avant que n’apparaissent les Amphibiens, et qu’une deuxième duplication a eu lieu beaucoup plus récemment, il y a environ 100 Ma.
Pistes d’exploitation pédagogique des données fournies
L’exploitation des données fournies sur les hormones antéhypophysaires et placentaires permet de dégager les notions liées aux mécanismes d’évolution du génome, et notamment à l’apparition de nouveaux gènes par duplication génique suivie de mutations indépendantes.
Ici, les mutations qui sont apparues ont amené à des fonctionnalités nouvelles pour les protéines codées puisque toutes ces hormones ont des rôles différents. Enfin, les gènes codant pour ces hormones étant situés sur des chromosomes différents, on illustre bien avec l’étude de cet exemple la souplesse du génome au cours de l’évolution.
Les gènes des hormones étudiées ici constituent deux familles multigéniques : la famille des gènes LH, FSH, TSH et HCG, et la famille des gènes de la GH, HPRL et HLP.
La comparaison des séquences protéiques et nucléiques au sein de chacun des groupes proposés (famille des gènes LH, FSH, TSH, HCG et famille des gènes GH, HPRL et HLP) permet de mettre en évidence des similitudes moléculaires et de dégager la notion de familles multigéniques. La notion de duplication génique suivie de mutations indépendantes permet alors d’expliquer l’apparition de nouveaux gènes à partir d’un même gène ancestral.
La prise en compte du taux d’identités permet de préciser les relations de parenté au sein de chacune de ces familles. Les données paléontologiques fournies permettent quant à elles de comprendre comment on peut dater approximativement certaines duplications géniques.
Enfin, les données fournies peuvent servir de support à une réflexion sur les effets des mutations et sur leur conservation :
-
l’étude des gènes et protéines LH et HCG permet de montrer comment une mutation peut, en supprimant un codon stop, entraîner un allongement de la protéine synthétisée, ce qui a contribué, avec l’apparition d’autres mutations, à l’obtention d’une protéine aux fonctionnalités nouvelles ;
-
l’alignement multiple des séquences protéiques des hormones LH, FSH, HCG et TSH permet de repérer l’existence de sites identiques remarquables, en particulier les cystéines. Or, ces résidus, en permettant la formation de ponts disulfures, jouent un rôle essentiel dans la stabilisation de la structure tridimensionnelle de la protéine et donc dans ses propriétés. On peut alors discuter du fait que ces sites soient très conservés d’une hormone à l’autre : une mutation à leur niveau doit arriver (aussi fréquemment qu’ailleurs dans la molécule), mais ne se fixe pas car elle entraîne une non-fonctionnalité de la protéine, donc nuit à la survie de l’individu porteur qui ne peut alors pas la transmettre.
Fichiers des séquences
-
GH-Genes-familleGH.edi : séquences nucléiques strictement codantes (du codon d’initiation au codon stop) des gènes de GH (hormone de croissance produite par l’antéhypophyse), HPRL (hormone prolactine produite par l’antéhypophyse), HLP (hormone lactogène placentaire produite par le placenta) ;
-
GH-Hormones-familleGH.edi : séquences protéiques des hormones GH (hormone de croissance produite par l’antéhypophyse), HPRL (hormone prolactine produite par l’antéhypophyse), HLP (hormone lactogène placentaire produite par le placenta).
-
LH-Genes-familleLH.edi : séquences nucléiques strictement codantes (du codon d’initiation au codon stop) des gènes de LH (hormone lutéinique produite par l’antéhypophyse), FSH (Folliculate Stimulating Hormon produite par l’antéhypophyse), TSH (Thyroïd Stimulating Hormon, produite par l’antéhypophyse) et HCG (hormone gonadotrophine chorionique, produite par les villosités choriales) ;
-
LH-Hormones-familleLH.edi : séquences protéiques des hormones LH (hormone lutéinique produite par l’antéhypophyse), FSH (Folliculate Stimulating Hormon produite par l’antéhypophyse), TSH (Thyroïd Stimulating Hormon, produite par l’antéhypophyse) et HCG (hormone gonadotrophine chorionique, produite par les villosités choriales).
Comparaisons obtenues pour la famille des gènes LH, FSH, TSH et HCG
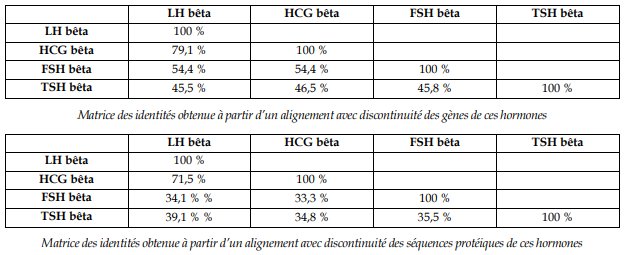
Le plus fort taux d’identité est entre les gènes LH bêta et HCG bêta (ou entre les hormones elles-mêmes). On en déduit que ces deux gènes dérivent d’un gène ancestral récent, par un phénomène de duplication génique ; les différences qu’ils présentent aujourd’hui s’expliquent par la fixation de mutations différentes dans les deux cas :
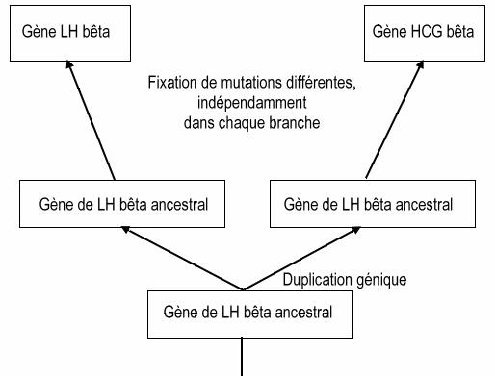
La LH bêta étant présente chez tous les Vertébrés alors que la HCG bêta n’étant présente que chez quelques ordres de Mammifères placentaires (Primates, Équidés), on peut dater approximativement la duplication à l’origine des gènes de ces deux hormones : elle est postérieure à l’apparition des Mammifères placentaires. Les Équidés n’étant pas les Mammifères les plus étroitement apparentés aux Primates, on peut supposer que l’apparition de HCG résulte d’une évolution convergente dans les lignées des Primates et des Équidés. D’autre part, cela permet de dire que le gène ancestral devait coder pour une molécule de type LH bêta.
La mise en relation des mutations dans les gènes LH bêta et HCG bêta avec les différences observées au niveau protéique permet de discuter de l’effet de certaines mutations : ici, une mutation par délétion en position 402 apparue dans le gène HCG bêta provoque la disparition d’un codon stop et donc l’allongement de la protéine synthétisée (la HCG bêta possède 165 acides aminés, la LH bêta n’en possède que 141).
Les taux d’identité entre les gènes FSH bêta, TSH bêta et LH bêta/HCG bêta sont suffisants pour dire que ces gènes sont homologues (et que ces protéines sont homologues), mais les différences ne sont pas assez significatives pour pouvoir réaliser une phylogénie pertinente (entre 54,4 % et 45,5 %, la différence n’est pas assez nette pour pouvoir vraiment distinguer sur une phylogénie les duplications à l’origine de ces gènes, d’un point de vue chronologique). Comme on obtient quatre gènes à partir d’un gène ancestral unique, on peut dire qu’il y a eu trois duplications dans cette famille de gènes.
Les gènes des hormones FSH et TSH n’étant pas sur le même chromosome que les gènes LH et HCG, il faut également faire intervenir un phénomène de transposition (ou translocation) de gènes lors de chacune des deux duplications les plus anciennes. Les quatre hormones n’ayant pas les mêmes rôles, on peut discuter de l’effet des mutations apparues et fixées différemment selon les branches ; ces mutations sont à l’origine des fonctionnalités nouvelles acquises par ces hormones.
Comparaisons obtenues pour la famille des gènes GH, HPRL et HLP
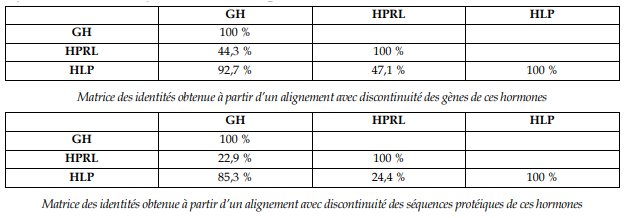
Le plus fort taux d’identité est entre les gènes GH et HLP (ou entre les hormones elles-mêmes). On en déduit que ces deux gènes dérivent d’un gène ancestral récent, par un phénomène de duplication génique ; les différences qu’ils présentent aujourd’hui s’expliquent par la fixation de mutations différentes dans les deux cas :
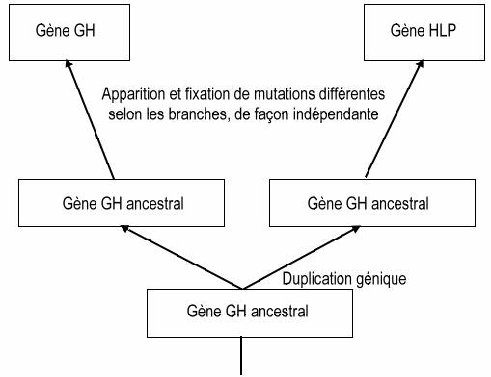
La GH étant présente chez tous les Vertébrés alors que la HLP n’est présente que chez les Mammifères placentaires, on peut dater approximativement la duplication à l’origine des gènes de ces deux hormones chez l’Homme : elle a eu lieu chez une espèce à l’origine de tous les placentaires actuels, soit environ entre 85 et 100 Ma. D’autre part, cela permet de dire que le gène ancestral devait coder pour une molécule de type GH.
Les taux d’identité entre les gènes HLP et GH/HPRL sont beaucoup plus faibles, mais suffisants pour dire que ces gènes sont homologues (et que ces protéines sont homologues), surtout si l’on considère la conservation de plusieurs acides aminés à des positions précises dans les protéines codées et la structure globale identique de ces gènes. Comme on obtient trois gènes à partir d’un gène ancestral unique, on peut dire qu’il y a eu deux duplications dans cette famille de gènes.
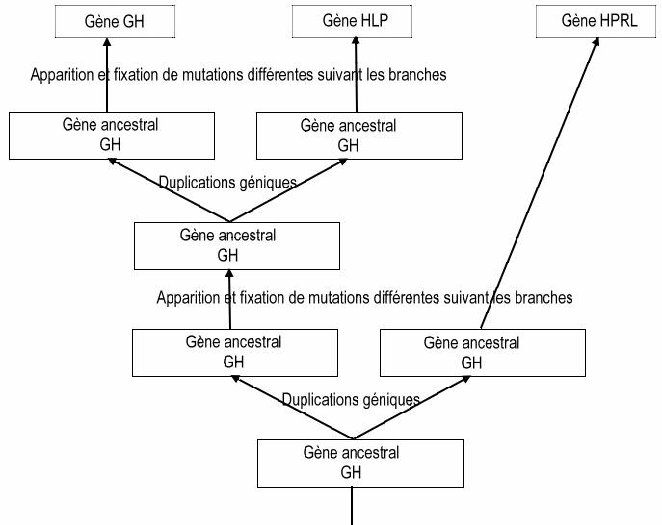
Les gènes des hormones GH et HLP n’étant pas sur le même chromosome que le gène HPRL, il faut également faire intervenir un phénomène de transposition (ou translocation) de gènes lors de la duplication la plus ancienne (tous les Vertébrés possédant les gènes GH et HPRL, on peut dire que la duplication à l’origine de ces deux gènes est plus ancienne que l’apparition du groupe des Amphibiens, soit il y a environ 400 Ma).
Les trois hormones n’ayant pas les mêmes rôles, on peut discuter de l’effet des mutations apparues et fixées différemment selon les branches ; ces mutations sont à l’origine des fonctionnalités nouvelles acquises par ces hormones.