
La différenciation sexuelle morphologique
Rédigée par Françoise Jauzein, ACCES 2003
Relue par Solange Magre, Université Paris 6, et Bernard Vigier, INRA
Révisé en 2018
Le stade de la gonade indifférenciée
Même si le sexe de l'embryon est déterminé dès la fécondation, la première ébauche de gonade n'apparaît, dans l'espèce humaine, qu'à la cinquième semaine de développement. Il s'agit d'une crête génitale formée à partir d'une prolifération du mésenchyme du mésonéphros (ou corps de Wolff ) et recouverte de l'épithélium coelomique.
Les cellules germinales primordiales ont une origine bien différente, elles proviennent d'une région de l'épiblaste adjacente à l'ectoderme extra-embryonnaire. En détectant leur activité phosphatase alcaline spécifique, on peut suivre leur cheminement (à partir de la quatrième semaine post-conception chez l'homme) de l'extrémité postérieure de la ligne primitive (à la base de l'allantoïde) à l'endoderme de l'intestin postérieur, puis le territoire gonadique (à la cinquième et sixième semaine) via le mésentère dorsal. Des expériences in vitro ont montré que les crêtes génitales exercent un chimiotactisme positif sur les cellules germinales primitives.
L'ébauche de gonade ainsi formée renferme des cellules germinales entourées de cellules somatiques. Des vaisseaux sanguins parcourent cette ébauche.
|
A. Jost, dans "La reproduction chez les mammifères et l'homme" Ellipses INRA |
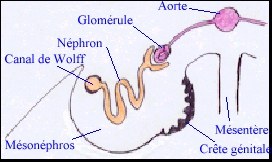
Stade indifférencié ou crête génitale apparue chez l'homme à la cinquième semaine La gonade se présente comme un épaississement de la paroi du mésonéphros. Chez la souris elle se compose d'un blastème de cellules germinales et somatiques (sans aucune organisation de type épithéliale) alors que chez l'homme on distingue des cordons épithéliaux et du tissu mésenchymateux. On pense actuellement que l'épithélium coelomique recouvrant la crête et le mésenchyme sous-jacent participent ensemble à l'édification de la partie somatique de la gonade. |
La différenciation gonadique
Chronologie de la différenciation des gonades
|
Principales étapes chez les mâles (M) et les femelles (F) |
Stade de développement chez la souris |
Stade de développement chez l'homme |
|
Migration des cellules génitales dans la crête génitale |
10 à 11 jpc |
4 à 5 semaines |
|
Différenciation des cordons séminifères (M) |
12 jpc |
7 semaines |
|
Différenciation des cellules de Leydig (M) |
13 jpc |
8 semaines |
|
Début de la prophase méiotique (F) |
13 à 14 jpc |
9 semaines |
|
Début de la folliculogénèse (F) |
3 jours après la naissance |
14 semaines |
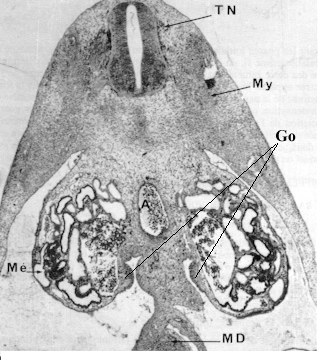
Dès la septième semaine de développement, chez les embryons humains de sexe génétique masculin (46, XY) , la gonade indifférenciée commence à se transformer en testicule, alors que chez les embryons de sexe génétique féminin (46 , XX), la transformation ne commence qu'à partir de la huitième semaine et les ovaires ne sont reconnaissables qu' à la dixième semaine grâce à la prophase méiotique de leurs cellules germinales.
Le contrôle de l'entrée en méiose des cellules germinales n'est pas totalement élucidé. Des cellules germinales mâles, égarées en position ectopique (dans le mésonéphros ou la surrénale) au cours de leur migration, entrent en prophase méiotique au même moment que les cellules germinales femelles. Ceci laisse à penser que les cellules germinales sont programmées pour entrer en meiose et que c'est l'environnement testiculaire qui bloque ce processus jusqu'à la puberté, période où la spermatogénèse se met en place.
|
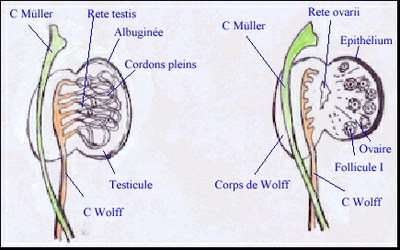
Elle est caractérisée par la formation de cordons séminifères, c'est à dire de cordons épithéliaux composés de cellules germinales (les spermatogonies, qui subissent une période de multiplication puis un arrêt mitotique dû à l'environnement testiculaire) et de cellules somatiques (les cellules de Sertoli). La différenciation des cellules de Sertoli (cytoplasme clair et volumineux, REG développé) est le premier évènement de l'organogénèse testiculaire. Ces cellules s'associent les unes aux autres par des interdigitations et des jonctions membranaires, englobant progressivement les cellules germinales et donnant ainsi naissance à des cordons séminifères pleins. En même temps se différencie la tunique albuginée, assise de tissu conjonctif se développant sous l'épithélium coelomique. Entre ces cordons, au sein du mésenchyme, se différencient, à partir de la huitième semaine, les cellules de Leydig (goutelettes lipidiques, REL) dont l'origine précise (mésonéphros ou crête) n'est pas encore établie. |
Après une phase de multiplication active (comme chez le mâle) les cellules germinales de la femelle entrent spontanément en prophase méiotique et se bloquent en ovocyte I au stade diplotène (ou dyctié). C'est le premier évènement de la différenciation femelle de la gonade. Les follicules, eux, se forment par regroupement de cellules somatiques avec des ovocytes, au sein de structures épithéliales, appelées cordons ovariens ou ovigères, qui restent en contact avec l'épithélium de surface. Lors du début de la folliculogénèse (qui a lieu chez certaines espèces après la naissance), ces cordons se fragmentent et chaque follicule primordial se constitue par association d'un ovocyte I et d'une assise de cellules somatiques, précurseurs des cellules de la granulosa. Les cellules thécales se différencient plus tard, autour des follicules en croissance. La présence de cellules germinales est indispensable à la formation des follicules car en leur absence, que ce soit pour des raisons génétique ou exogène, des cordons stériles se mettent en place mais dégénèrent ensuite, laissant un ovaire composé uniquement de stroma. |


Pendant la vie foetale et la petite enfance ces cordons restent pleins, ils se creusent en tubes séminifères au moment de la puberté, on observe alors la jonction entre ces tubes, le reste des tubules mésonéphrotiques et le canal de Wolff qui fonctionnera en spermiducte. |
|
Les connexions uro-génitales dégénèrent en même temps que le mésonéphros (laissant parfois quelques formations kystiques comme le rete ovarii) |
L'évolution du tractus
Au moment où débute la différenciation gonadique mâle, le tractus est encore sexuellement indifférencié.
Le mésonéphros s'est mis en place, pendant la quatrième semaine du développement foetal humain, à partir du mésoblaste situé de la cinquième paire de somites cervicaux à la quatrième paire de somites lombaires. Dans le mésonéphros, des néphrons débouchent dans le canal de Wolff, qui, à la cinquième semaine, continue son extension sous la vessie jusqu'au sinus-urogénital, et forme un diverticule à la hauteur de l'ébauche du métanéphros (futur rein). Ce diverticule donnera l'uretère, son orifice se séparera du canal de Wolff lors de la croissance du sinus-urogénital (le sinus uro-génital s'étend de la base de la vessie jusqu'à l'orifice uro-génital, situé au niveau du périnée, sous le tubercule génital qu'encadrent les bourrelets génitaux).
Quant au canal de Müller, il est issu d'une invagination de l'épithélium coelomique de la région antérieure du mésonéphros, ayant pris naissance à proximité de la région antérieure du canal de Wolff au cours de la sixième semaine, pendant la période de formation des crêtes génitales. Il se développe ensuite vers l'avant et vers l'arrière jusqu'au sinus uro-génital en cheminant le long du canal de Wolff.
 Cavité abdominale d'un foetus humain de 7 semaines environ Dans "la reproduction chez les mammifères et l'homme". Ellipses, INRA |
|

La différence essentielle entre les deux sexes porte alors sur le devenir des canaux de Wolff et de Müller.
Chronologie de la différenciation du tractus génital
|
Principales étapes |
Age du foetus murin |
Age du foetus humain |
|
Régression des canaux de Müller |
14 jpc |
8 semaines |
|
Masculinisation des organes génitaux externes, distance ano-génitale augmentée (M) |
16 jpc |
9 à 10 semaines |
|
Vésicules séminales (M) |
16 jpc |
10 semaines |
|
Bourgeons prostatiques (M) |
17 jpc |
10 sem |
|
Régression des canaux de Wolff (F) |
15 jpc |
10 sem |
|
Fin de la migration testiculaire (M) |
18 jpc |
12 semaines |
|
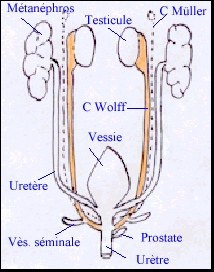
Chez le foetus mâle les canaux de Müller régressent totalement, ne laissant qu'un reliquat facultatif au niveau de l'épididyme (l'hydatide sessile) et un petit diverticule de l'urètre prostatique (l'utricule prostatique). Dans leurs partie supérieure les canaux de Wolff régressent en ne laissant qu'un reliquat (l'hydatide pédiculée). Dans leur partie moyenne, ils constituent les canaux épididymaires, où s'abouchent les canaux efférents (anciens tubules mésonéphrétiques entrés en communication avec le rete testis). Dans leur portion inférieure ils deviennent les canaux éjaculateurs qui débouchent dans l'urètre prostatique. A l'union entre ces segments, deux bourgeonnements de la paroi forment les ébauches des vésicules séminales. La partie distale des conduits génitaux et urinaire dépend de l'évolution du sinus uro-génital, essentiellement à partir de la neuvième semaine. Les organes génitaux externes se forment de la neuvième à la quatorzième semaineà partir du tubercule génital. Les orifices génital et urinaire sont communs. |
Chez le foetus femelle les canaux de Wolff régressent entièrement en laissant quelques reliquats embryonnaires sous la forme de petits kystes allongés (canaux de Gärtner) que l'on retrouve contre les parois latérales de l'utérus et du vagin et, dans leurs parties hautes, l'hydatide pédiculée et l'époophore. Les canaux de Müller forment, dans leur segments supérieurs, les trompes de Fallope. Dans leurs segments inférieurs, ils fusionnent pour constituer le canal utéro-vaginal (présentant un renflement postérieur, le tubercule de Müller) alors que les segments moyens évoluent en deux cornes utérines. A partir du quatrième mois, les deux cornes utérines se soudent en un utérus unique, le tubercule de Müller devient le col de l'utérus et le vagin se cavite. La partie distale des conduits génital (partie basse du vagin) et urinaire dépend de l'évolution du sinus uro-génital, à partir du quatrième mois. Les organes génitaux externes se forment au cours du troisième mois à partir du tubercule génital. Les orifices urinaire et génital sont totalement distincts l'un de l'autre. |

Enfin, la migration des testicules en dehors de l'abdomen, dans le scrotum, se fera durant le septième et le huitième mois de la grossesse. La position des testicules est déterminée par deux ligaments génitaux: le ligament suspenseur crânial et le gubernaculum (ou ligament génital caudal). Chez les mâles, c'est la croissance des gubernaculums qui permet la descente des testicules en position scrotale. L'absence de cette descente testiculaire, ou cryptorchidie, entraîne une stérilité.