
L’ADN mitochondrial et l’origine d’Homo sapiens
L’ADN mitochondrial et l’origine d’Homo sapiens
Jean-Claude Hervé
I - Bref historique sur l’enseignement de l’origine de Homo sapiens.
II - Les données paléontologiques sur l’origine des Hommes modernes
-
Quelques repères sur les Homo des deux derniers millions d’années
-
L’ADN mitochondrial des Hommes actuels et l’origine d’Homo sapiens
I - Bref historique sur l’enseignement de l’origine de Homo sapiens
L’étude de la lignée humaine est inscrite dans les trois derniers programmes de terminale. Dans le programme de la fin des années 80 et aussi dans celui appliqué pour la première fois en septembre 1994, cette étude est envisagée sous le paradigme aujourd’hui dépassé de l’hominisation : « Les caractéristiques morphologiques, anatomiques, culturelles qui distinguent l’Homme se sont mises en place à travers l’individualisation rapide de formes humaines : certains Australopithèques, Homo habilis, Homo erectus, Homo sapiens. On est encore à une phylogénie de type gradiste avec la reconnaissance du groupe des grands singes, d’où serait issue, par des transformations progressives, la lignée humaine . Suivant cette conception d’une évolution linéaire, les fossiles de cette lignée ne sont que des intermédiaires entre le grand Singe et l’Homme. Dans cette perspective, l’origine des Homo sapiens n’est pas abordée, l’Homo erectus étant implicitement l’ancêtre d’Homo sapiens.
Avec le programme de 2002, la perspective change nettement puisque les relations de parenté entre les êtres vivants sont abordées suivant une démarche cladiste et qu’on cible dans l’étude de la lignée humaine sur son caractère buissonnant. Pour la première fois, est inscrite au programme l’étude de l’origine des Hommes modernes, Homo sapiens. On privilégie une des hypothèses en utilisant toutefois le conditionnel : « Homo sapiens serait une nouvelle espèce apparue en Afrique ou au Proche Orient il y a 100.000 à 200.000 ans et aurait colonisé tous les continents en remplaçant Homo erectus ». La seule argumentation fournie pour asseoir cette affirmation est la proximité génétique des populations humaines, partageant toutes les mêmes allèles des gènes, mais avec une fréquence variable. Dans l’accompagnement du programme de terminale S, on dit d’ailleurs que ne sont pas exigibles la discussion sur l’origine polycentrique ou monocentrique de l’Homme moderne, et les arguments liés aux données sur l’ADN mitochondrial. Notons toutefois que des questions de type « pratique du raisonnement scientifique » posées au bac ont fait appel à des données sur l’ADN mitochondrial dans la mesure où elles n’impliquaient que des types de raisonnement que les élèves devaient maîtriser.
Or, à l’époque où est paru ce programme, la controverse était encore forte entre deux théories, l’une dite multirégionale, l’autre « Out of Africa » sur l’origine des Hommes modernes. Pour cette question comme pour beaucoup d’autres relatives à l’enseignement de la lignée humaine (par exemple celle des caractéristiques de l’ancêtre commun Homme- Chimpanzé) il est de loin préférable de ne pas vouloir enseigner des vérités mais d’aborder les modèles explicatifs retenus par les chercheurs confrontés avec les données dont on dispose. Autrement dit l’enseignement de la lignée humaine est un très bon support pour faire saisir comment fonctionne la science.
Les théories sur l’origine des Hommes modernes ont d’abord été émises à partir des donnés paléontologiques et archéologiques. A partir de 1986 et l’article de Rebecca Cann et Allan Wilson intitulé : « Mitochondrial DNA and human evolution » paru dans Science, les données génétiques relatives à l’ADN mitochondrial sont entrées dans le débat. Les auteurs comparaient les ADN mitochondriaux d’Africains, d’Asiatiques, de Caucasiens, d’Aborigènes australiens, de Nouvelle Guinée et en conclusion disaient que leurs travaux confirmaient l’origine unique africaine d’Homo sapiens. Les travaux de Cann et Wilson reposaient sur la comparaison des fragments obtenus à partir de l’action d’une douzaine d’enzymes de restriction sur un fragment d’ADN mitochondrial représentant environ 9% de la molécule complète. Depuis, près de 5000 génomes d’ADN mitochondrial complet ont été séquencés, appartenant à des individus de toutes les populations mondiales (le génome mitochondrial comprend près de 16600 paires de bases). Autrement dit, les données disponibles en 2010 permettent d’établir une phylogénie solide des ADN mitochondriaux des populations humaines permettant de tester les théories sur l’origine des Hommes modernes.
En outre, en 2008 et 2009 ont été publiées les séquences complètes d’ADN mitochondrial de 6 Néandertaliens, permettant la comparaison avec celles des Hommes modernes et la discussion sur la possibilité d’hybridation ou non entre Homo sapiens et Homo neanderthalensis.
Enfin, en 2010, a été publiée la séquence complète de l’ADN mitochondrial extrait d’une phalange d’un Homininé trouvé à Denisova , dans une grotte de Sibérie, ADN mitochondrial qu’on a comparé à ceux des Homo sapiens et des Néandertaliens. A partir de cette seule séquence, on a parfois affirmé la découverte d’une autre espèce d’Homo ayant vécu en même temps que Néandertal et Homo sapiens !
Bref, les données sur l’ADN mitochondrial se sont considérablement enrichies durant ces 10 dernières années et ont pris beaucoup d’importance vis-à-vis de la compréhension de l’histoire récente de la lignée humaine.
II - Les données paléontologiques sur l’origine des Hommes modernes
1 – Quelques repères sur les Homo des deux derniers millions d’années
Afin de bien saisir les théories sur l’origine des Hommes modernes, il est nécessaire d’avoir une vue d’ensemble sur les peuplements d’Homo dans les diverses régions du monde depuis l’apparition d’Homo erectus en Afrique, il y a deux millions d’années environ, puisque ces théories doivent rendre compte de ces peuplements.
Afrique
De 2 à 1 millions d’années, on connaît plusieurs sites ayant livré des ossements d’Homo erectus, avec une industrie acheuléenne caractérisée par les bifaces. A cause de différences au niveau du crâne entre les erectus africains et les erectus asiatiques, on donne parfois le nom d’Homo ergaster aux erectus africains, réservant l’appellation d’erectus aux spécimens asiatiques. A partir de 1 million d’années, les Homo erectus africains sont remplacés par des formes ayant une capacité cérébrale plus importante et dont les proportions crâniennes se rapprochent davantage de celles de l’Homme moderne. On donne parfois le nom d’Homo heidelbergensis à ces homininés datés de 600.000 ans environ. A partir de 500.000 ans, on a trouvé des fossiles aux caractéristiques crâniennes se rapprochant de celles des Hommes modernes, tout en conservant certains états primitifs. Les premiers Hommes modernes trouvés en Afrique remontent à près de 200.000 ans. C’est ainsi qu’en juin 2003, un article paru dans la revue Nature citait trois crânes trouvés en Ethiopie, présentant les caractéristiques des Hommes modernes. La vision d’ensemble qui se dégage des fossiles africains, certes fragmentaires, est celle d’une série plus ou moins continue allant des premiers Homo erectus aux Hommes actuels.
Asie
Les premiers Homo ayant quitté l’Afrique ont été trouvés à Dmanisi en Géorgie, aux portes de l’Europe et de l’Asie. Ils sont datés de 1,8 millions d’années. Par rapport aux Homo erectus africains, ils présentent des états primitifs (notamment une faible capacité cérabrale) de sorte qu’on a parfois créé une espèce particulière, Homo georgicus pour ces spécimens.
Les Homo erectus sont bien connus en Indonésie à Java (les premiers découverts par Dubois à la fin du 19ème siècle ont été initialement appelés Pithecantropus erectus) et en Chine, près de Pékin (appelés pendant longtemps Sinanthropus pekinensis). Les méthodes modernes de datation absolue ont permis de dater récemment divers spécimens. Ainsi des fossiles trouvés à Java, à Mojokerro et à Sangiran, ont été datés de 1,8 et 1,6 millions d’années environ. Ils témoignent donc de l’ancienneté du peuplement de l’Asie par les Homo erectus. En revanche, d’autres Homo erectus de Java, ceux de Ngandong, ont été datés de 53.000 à 27.000 ans. Ils possédaient toutefois une capacité crânienne plus forte que celle des premiers Homo erectus de sorte qu’on les appelle parfois Homme de Solo.
Les premiers fossiles d’Hommes modernes en Asie ont été récemment découverts. En 2007, des fragments du squelette d’Hommes modernes ont été découverts en Chine, près de Pékin, et ont été datés de 38.000 ans. Enfin, on connaît en Asie des spécimens qui n’entrent pas dans la variation morphologique des Homo erectus et qui ne sont pas des Hommes modernes. C’est le cas des fossiles de Dali qui combinent des caractères primitifs avec des traits d’Homo sapiens, appartenant à une fourchette de temps allant de 200.000 ans à 100.000 ans.
Europe
Les plus anciens fossiles d’Homo en Europe ont été trouvés en Espagne, et ils sont datés de 800.000 ans (sites de la Gran Dolina d’Atapuerca). Les découvreurs ont créé pour ces fossiles une nouvelle espèce, Homo antecessor, loin d’être reconnue par la communauté des paléoanthropologues. On peut les considérer comme des Homo erectus européens. En tout cas ils ne possèdent pas d’affinités néandertaliennes. Des données archéologiques laissent même à penser que les Homo ont pénétré dans l’Europe du sud depuis plus de 1 millions d’années, mais l’industrie associée est primitive ne comportant pas de bifaces.
Le second gisement de fossiles européens, celui nommé Sima de los Huesos, est aussi en Espagne (et proche du gisement précédent) et daté de 500.000 ans environ. Ces fossiles ressemblent aux homo Heideilbergensis africains mais avec déjà certains caractères dérivés des néandertaliens. A partir de cette époque, on a trouvé dans toute l’Europe des fossiles présentant une mosaïque variable d’états dérivés de néandertaliens, qu’on range souvent comme prénéerdantaliens. Les premiers fossiles avec toutes les caractéristiques morphologiques, notamment crâniennes, définissant les néandertaliens sont datés de 130.000 ans environ. Les néandertaliens persistent en Europe jusqu’à 30.000 ans.
Les premiers Homo sapiens d’Europe, les Hommes de Cro-Magnon, sont datés de 40.000 ans ; ils ont donc cohabité avec les néandertaliens pendant une dizaine de milliers d’années.
Autres régions
- Le proche orient est remarquable par l’existence de sites ayant livré à la fois des fossiles d’Homo sapiens et de néandertaliens. C’est le cas par exemple du site de la grotte de Qafzeh (Israël) dont les fossiles sont datés de 100.000 ans environ.
- Les premiers Homo reconnus en Australie sont des Homo sapiens datés de 50.000 ans environ.
- De même, l’installation des Homo dans les Amériques a été le fait d’Homo sapiens mais plus tardivement qu’en Australie (12.000 ans pour l’hypothèse courte, 30.000 ans pour l’hypothèse longue).
2 – Les théories sur l’origine des Hommes modernes
A grands traits, on peut dire que deux théories se sont affrontées pour expliquer l’origine des Homo sapiens modernes.
L’une dite théorie multirégionale est basée sur la migration des Homo erectus hors d’Afrique à partir de 2 millions d’années environ. Elle propose que les populations ancestrales d’Homo erectus répandues à travers le monde ont graduellement et indépendamment évolué pour donner d’abord des Homo sapiens archaïques, puis pleinement l’Homme moderne. Ce modèle envisage donc l’apparition d’Homo sapiens en de multiples endroits de la planète. Dans ce cadre, les néandertaliens sont considérés comme la version européenne des sapiens archaïques. L’une des conséquences de ce modèle est que les populations actuelles géographiquement distinctes auraient des racines très lointaines. Comme les populations des différentes parties du monde étaient séparées par de grandes distances et étaient soumises à différents types de conditions environnementales, il y a eu chez elles une variation régionale de morphologie.
La deuxième théorie dite « Out of Africa » admet une origine unique de tous les Hommes modernes, leur apparition en un lieu précis, suivi d’une migration dans toutes les régions du monde. En accord avec les données paléontologiques qui révèlent que les plus anciens fossiles d’Homo sapiens ont été trouvés en Afrique (en Ethiopie avec une datation de 170.000 ans environ), l’Homme moderne apparu en Afrique, il y a 200.000 ans environ s’est ensuite dispersé en Afrique en de nombreuses populations. Puis, vers 50.000 ans, les individus d’une de ces populations régionales ont quitté l’Afrique et se sont dispersés à travers le monde en remplaçant les autres Homo déjà présents. Ainsi, les Hommes modernes en Europe ont fini par remplacer les néandertaliens après une à deux dizaines d’années de cohabitation. Dans la version la plus extrême de cette théorie de « Out of Africa » il n’y a pas eu d’hybridation des Homo sapiens avec les populations d’Homo déjà présentes dans les autres régions du monde. Cette théorie met donc en relief l’origine récente et commune de tous les Hommes actuels.
Il est bien sûr possible de discuter de la validité de ces deux théories par rapport aux données fossiles. Par exemple, une étude comparative un peu poussée de l’anatomie des néandertaliens et des Hommes modernes rend très peu probable une évolution continue ayant mené des néandertaliens aux hommes modernes. Nous allons seulement envisager comment les données génétiques acquises durant les 20 dernières années et exploitables avec Phylogène permettent de tester les théories sur l’origine des Hommes modernes.
3 - L’ADN mitochondrial des Hommes actuels et l’origine d’Homo sapiens
On ne peut, bien sûr, avec Phylogène exploiter les séquences d’ADN mitochondrial des quelques 5000 individus actuellement référencés. Nous avons extrait une banque de 53 ADN mitochondriaux complets provenant d’individus de différents endroits du globe. Ce sont les 53 séquences du premier article ayant fait une synthèse sur les ADN mitochondriaux complets des Hommes actuels, article publié dans la revue Nature en 2000 (Mitochondrial genome variation and the origin of modern humans. Ingman et al).
La figure ci-dessous illustre l’arbre phylogénétique des ADN mitochondriaux des 53 individus obtenu avec Phylogène en utilisant la méthode UPGMA.
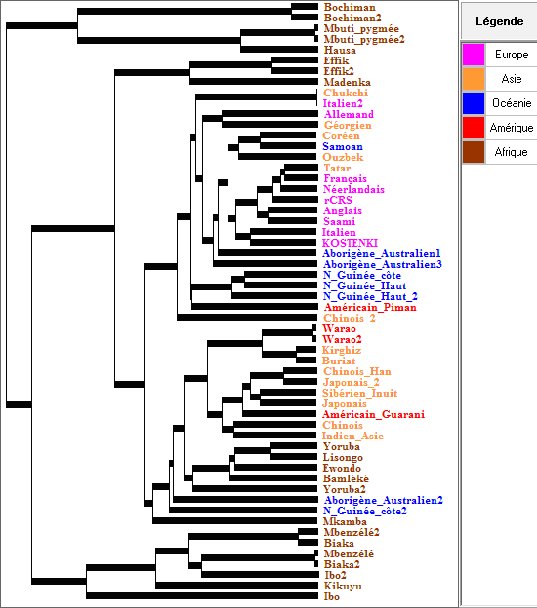
Cet arbre a été obtenu en utilisant la méthode phénétique la plus basale (la méthode UPGMA) dont le principe peut être compris facilement par les lycéens, méthode qui repose sur la prise en compte du nombre de différences entre les diverses séquences prises deux à deux. Néanmoins dans ses grandes lignes, cet arbre fournit les mêmes informations que celui obtenu par la méthode plus sophistiquée du Neighbor joining.
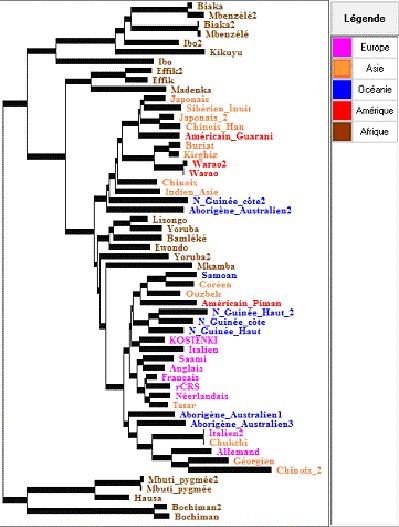
Les points les plus fondamentaux de cet arbre ne sont pas contredits par les arbres cladistes plus récents obtenus en considérant les divers sites variables de l’ADN mitochondrial. Néanmoins, il ne faut pas faire dire à cet arbre obtenu avec Phylogène plus que ce qu’il indique.
En premier lieu, on voit qu’à partir de la racine de l’arbre les 3 branches qui se détachent en premier sont exclusivement africaines, ce qui est en accord avec un peuplement initialement africain de Homo sapiens. Ensuite, on constate que toutes les populations non africaines se rassemblent en un nœud mais que de ce grand ensemble, font aussi partie certaines populations africaines (Yoruba, Lisongo, Ewondo, Bamleke, Mkamba). Cela est accord avec l’hypothèse de l’origine africaine d’Homo sapiens : ces africains pourraient être les descendants de la population africaine qui a émigré à l’origine et peuplé le reste du monde.
Une autre façon d’exploiter ces données est d’évaluer l’importance des différences entre les ADN mitochondriaux des africains d’une part, des non africains d’autre part. On peut utiliser pour cela, la collection sur les africains seulement, et celle sur les non africains. On constate que les différences entre les ADN mitochondriaux des diverses populations africaines sont beaucoup plus importantes qu’entre les ADN mitochondriaux des autres populations mondiales. Si on admet le principe de l’horloge moléculaire suivant lequel le rythme des mutations dans les diverses populations a été le même, cette grande variabilité des africains indique une période d’évolution plus longue : les premières populations humaines étaient africaines, ce qui réfute l’hypothèse multirégionale.
Il serait toutefois hasardeux d’aller plus loin dans l’exploitation de cet arbre et de vouloir y rechercher des indications sur l’histoire des populations non africaines. Il faut pour cela une analyse plus fine utilisant le repérage de sites caractéristiques dans les diverses populations, (c’est une branche d’étude très riche actuellement).
L’exploitation de cet arbre conduit non seulement à valider l’hypothèse de l’origine africaine des Hommes modernes, mais encore à l’affiner un peu. Il suggère qu’Homo sapiens est apparu en Afrique (dans l’Afrique sub-saharienne d’après les populations africaines aux ADN mitochondriaux les plus éloignés des autres. A partir de cette souche initiale, il y a eu une dispersion en Afrique avec individualisation de populations. Les individus d’une des populations africaines se sont dispersés et ont été à l’origine des populations des autres régions du monde par la suite.
4 - L’ADN mitochondrial des néandertaliens et des sapiens
Les seules données sur l’ADN mitochondrial des sapiens actuels corroborent l’origine africaine d’Homo sapiens. Cela n’exclut pas cependant la possibilité d’hybridation entre les populations de Homo sapiens ayant quitté l’Afrique et les Homo déjà présents dans les diverses régions du monde. En particulier, en Europe, y a–t’il eu métissage entre sapiens et néerdantaliens étant donné que les deux ont cohabité pendant une dizaine de milliers d’années ?
Les deux collections sur les séquences complètes de l’ADN mitochondrial de fossiles de la lignée humaine, permettent de discuter de cette question. La première collection comprend les séquences complètes de 6 néandertaliens (séquences parues en 2008 et 2009), de plusieurs sapiens dont les africains Hausa et San, ainsi que celle de l’Homme de Kostenki, sapiens trouvé en Russie et daté de 30.000 ans environ. Les deux collections comprennent en plus la séquence mitochondriale de Denisova., séquence obtenue à partir de l’ADN mitochondrial extrait d’une phalange d’un doigt d’un Homininé , trouvé à Denisova, dans une grotte de Sibérie et datée de 30.000 à 50.000 ans.
Dans les deux collections, les séquences d’ADN mitochondrial des Chimpanzés servent d’outgroup. La deuxième s’en distingue par les séquences des diverses sous espèces de Chimpanzés communs, ce qui permet une discussion des différences constatées entre néandertaliens et sapiens.
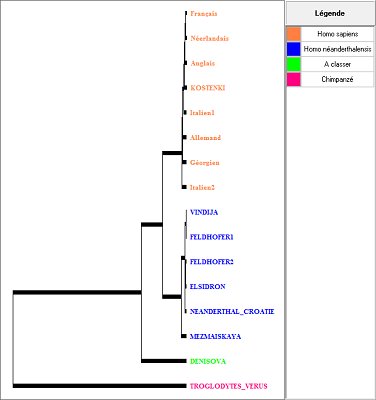
L’aspect le plus frappant de cet arbre est que toutes les séquences d’ADN mitochondrial néandertaliennes sont très proches les unes des autres et également distantes de toutes les séquences des sapiens, y compris de celle de Kostenki. L’examen de la matrice des distances montre qu’en gros les séquences néerdantaliennes diffèrent de celles de sapiens par 200 nucléotides environ. alors que la différence maximale entre les séquences de sapiens est de 100. Les séquences néerdantaliennes ne montrent pas de similitude plus forte avec celles des européens qu’avec celles des autres régions du monde. Cela indique qu’on ne trouve dans aucune des séquences d’ADN mitochondrial des sapiens jusqu’ici déterminées, un héritage de l’ADN mitochondrial d’une femme néerdantalienne. L’ADN mitochondrial n’apporte pas de preuve d’hybridation entre sapiens et néerdantaliens.
En appliquant le principe de l’horloge moléculaire, calibrée avec les séquences des chimpanzés, les chercheurs fournissent, à partir de ces données génétiques, une date de 600.000 ans environ pour la divergence des deux lignées, celle menant aux néandertaliens, et celle menant aux sapiens actuels, ce qui est en gros en accord avec les données paléontologiques. Quant à l’apparition d’Homo sapiens, ils la datent à 170.000-200.000 ans environ et la sortie d’Afrique à 60.000-50.000 ans environ.
Ces données suffisent-elles pour affirmer qu’Homo sapiens et Homo néanderthalensis appartiennent à deux espèces distinctes ?
L’importance des différences entre l’ADN mitochondrial de sapiens et celui de néandertal suffit-elle pour dire qu’il s’agit de deux espèces distinctes ? Il faudrait avoir un critère disant à partir de quel taux de différences on peut parler d’espèces distinctes. Les données sur les Chimpanzés permettent d’approcher le problème de façon critique. Le genre Pan est en effet représenté par le Chimpanzé commun (Pan troglodytes) qui comprend 4 sous espèces, et le Chimpanzé Bonobo. En évaluant le pourcentage de différence entre les deux espèces de Chimpanzés et en le comparant avec le pourcentage de variations intraspécifiques chez les Chimpanzés, on peut estimer un pourcentage de divergence significatif d’une spéciation. C’est ce que permet la deuxième collection sur les données génétiques de la lignée humaine.
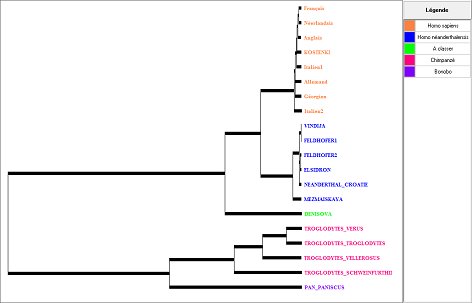
La seule observation de cet arbre montre que le pourcentage de différences entre les ADN mitochondriaux des deux espèces de Pan est nettement supérieur à celui entre sapiens et néandertal (l’exploitation de la matrice indique 4,07% de différences contre 1,27%). En outre, les différences intra spécifiques entre les espèces de Pan troglodytes sont aussi supérieures à celle des néandertaliens par rapport aux sapiens. Si on prend les données sur les Chimpanzés comme critères, on voit qu’à partir des seules donnés sur l’ADN mitochondrial, on ne peut conclure que Homo neanderthalensis et Homo sapiens appartiennent à deux espèces distinctes. (Bien sûr ce raisonnement suppose que l’ADN mitochondrial évolue avec la même vitesse chez les chimpanzés et les Homo).
Cela est confirmé par un article paru dans la revue Science en mai 2010 qui analyse la séquence de l’ADN nucléaire obtenue à partir de 3 néandertaliens et la compare avec celle de 5 Hommes modernes : un africain du sud (San), un Yoruba d’Afrique de l’ouest, un papou asien de Nouvelle Guinée, un Chinois et un Français. Les chercheurs ont trouvé des zones où l’ADN nucléaire des néandertaliens présente des caractéristiques retrouvées chez l’individu de nouvelle guinée, le Chinois et le Français mais pas chez les africains. Cela les conduit à penser qu’une partie (faible 4%) du génome des néandertaliens s’est recombinée avec celui d’homo sapiens hors d’Afrique. Ce qui suggère que des croisements ont eu lieu même s’ils ont été limités. En outre comme les caractéristiques néerdantaliennes sont présentes aussi bien chez le papou de nouvelle Guinée que chez le Chinois et le Français, on estime que l’hybridation a eu lieu avant que l’homme moderne n’atteigne l’Eurasie, probablement au Moyen orient où les néandertaliens sont présents de 100.000 ans à 40.000 ans.
Il faut bien voir que les auteurs de cet article (Green et al) sont les mêmes que ceux qui avaient obtenu les séquences d’ADN mitochondrial des néandertaliens en 2008 et 2009 et concluaient plutôt en sens inverse. Bon exemple de la façon dont fonctionne la science.
5 - Une nouvelle espèce d’Homo identifiée par la génétique ?
L’os de la grotte de Denisova dans des sédiments datés de 50.000 à 30.000 années, a été découvert près de lieux où on a trouvé des restes de néandertaliens et de sapiens. La collection des ADN mitochondriaux de la lignée humaine situe l’ADN de Denisova dans cette lignée. On s’aperçoit que cet ADN mitochondrial se situe tout à fait en dehors de la variabilité des homo sapiens et des néerdantaliens. Il présente environ deux fois plus de différences avec Homo sapiens que néandertal et il est également distant des sapiens que des néandertaliens. Manifestement, il n’appartient pas à ces deux groupes et en appliquant le principe de l’horloge moléculaire, on estime que la séparation de la lignée de Denisova avec celles des néerdantaliens et des sapiens remonte à un million d’années environ. Cela suggère une migration d’Homo hors d’Afrique imprévue. Faut-il pour autant affirmer qu’il s’agit d’une nouvelle espèce ? En utilisant le raisonnement vu précédemment, on constate que la différence entre l’ADN mitochondrial de Denisova par rapport à celle de sapiens et néandertal, est légèrement supérieure seulement à celle trouvée entre les sous espèces de chimpanzés et nettement inférieure à celle trouvée entre les deux espèces de Pan. Cela interdit de conclure trop hâtivement à la découverte d’une nouvelle espèce comme les médias s’en sont fait l’écho contrairement aux chercheurs qui sont restés plus prudents.