1. Le déterminisme du sexe chez les reptiles
Détermination du sexe par la température chez les reptiles
Lien vers le document original
Rédigé par Françoise Jauzein,
Relue par Claude PIEAU, Institut Jacques Monod

Différentes modalités de détermination du sexe chez les reptiles
Comme les poissons et les amphibiens, les reptiles présentent différents types de déterminisme du sexe.
Une détermination génotypique (GSD pour "genotypic" ou CSD pour "chromosomal sex determination"), contrôlée par un ou des gènes portés par les chromosomes sexuels, se rencontre chez tous les serpents, de nombreux lézards et une minorité de tortues. Chez les serpents, l'hétérogamétie est femelle, de type ZW/ZZ, alors que les deux mécanismes, hétérogamétie femelle ZW/ZZ et mâle XY/XX, se rencontrent chez les lézards et les tortues.
Chez d'autres espèces de reptiles, toutes ovipares, on peut rencontrer un déterminisme du sexe d'un type différent; la différenciation sexuelle des gonades dépend, chez elles, de la température d'incubation des oeufs (TSD pour "temperature dependant sex determination"). Les premières observations de ce phénomène datent de 1966 et furent réalisées sur un lézard: Agama agama.
Cette TSD est connue aujourd'hui chez tous les crocodiles déjà étudiés, la plupart des tortues, quelques lézards et les deux espèces de Sphenodon.
|
|
|
Différentes modalités de détermination du sexe par la température En haut, il n'existe qu'une seule température pivot et une seule gamme de température transitionnelle, entre températures masculinisantes et températures féminisantes, détermine le sexe. Le cas présenté correspond à celui rencontré chez de la majorité des tortues. En bas, deux températures pivot et deux gammes de températures transitionnelles déterminent le sexe. C'est le cas des crocodiles et de certaines espèces de tortues et de lézards. |

Chez ces espèces il existe des températures masculinisantes (TM) donnant 100% ou une majorité de mâles, des températures féminisantes (TF) donnant 100% ou une majorité de femelles et une gamme de températures de transition (TRT pour "transition range of température") qui fournit des mâles, des femelles et quelquefois des intersexués.Trois mécanismes de détermination thermique du sexe sont connus.
Chez de nombreuses tortues, les températures d'incubation inférieures à la TRT fournissent des mâles et celles qui sont supérieures à cette TRT donnent des femelles. Chez quelques lézards et les Sphenodon, c'est l'inverse. Cependant, chez d'autres espèces de tortues, de lézard et chez les crocodiles, il existe deux TRT. Les températures situées entre ces deux TRT fournissent des mâles, alors que les températures situées en dehors de cette gamme intermédiaire, plus basses ou plus hautes, donnent des femelles. Au sein d'une TRT, la température "pivot" est celle qui détermine un sex-ratio de 1 (50% de mâles et 50% de femelles).
Les deux types de détermination du sexe présentant une seule gamme de températures de transition pourraient correspondre à une évolution à partir d'un modèle ancestral comportant deux gammes transitionnelles (modèle qui se retrouve dans les trois groupes de reptiles présentant un déterminisme du sexe par la température), le modèle que l'on rencontre chez la plupart des tortues correspondant à une conservation de la gamme TRT2 seulement et celui, inverse, que l'on rencontre chez les lézards et les Sphenodon, correspondant à la conservation de la gamme TRT1.
Période de sensibilité à la température
| La période thermosensible du développement embryonnaire est connue pour quelques espèces de crocodile, de tortue et un lézard. Elle représente 18 à 30% de la durée du développement embryonnaire et correspond aux premiers stades de la différenciation des gonades. Les études précédentes ont été obtenues en maintenant des oeufs à température constante, alors qu'en réalité, dans le nid, la température d'incubation dépend, en milieu tempéré et si les nids sont peu profonds (espèces de tortues de petite taille) de l'heure de la journée et du temps. Chez les grosses tortues marines qui font, en milieu tropical, des nids profonds, les variations de température nycthémérales sont peu importantes. Cependant, en Guyane par exemple, les tortues marines pondent jusqu'à une dizaine de fois, de février à septembre. Durant cette période la température dans le nid sera plus importante pendant la saison sèche. Les pourcentages de mâles et de femelles observés au moment de l'éclosion dépendent, au cours de la période de thermosensibilité, des fractions du développement passées en dessous ou au dessus de la TRT. |
 |
La période thermosensible du développement embryonnaire est connue pour quelques espèces de crocodile, de tortue et un lézard. Elle représente 18 à 30% de la durée du développement embryonnaire et correspond aux premiers stades de la différenciation des gonades.
Les études précédentes ont été obtenues en maintenant des oeufs à température constante, alors qu'en réalité, dans le nid, la température d'incubation dépend, en milieu tempéré et si les nids sont peu profonds (espèces de tortues de petite taille) de l'heure de la journée et du temps. Chez les grosses tortues marines qui font, en milieu tropical, des nids profonds, les variations de température nycthémérales sont peu importantes. Cependant, en Guyane par exemple, les tortues marines pondent jusqu'à une dizaine de fois, de février à septembre. Durant cette période la température dans le nid sera plus importante pendant la saison sèche.
Les pourcentages de mâles et de femelles observés au moment de l'éclosion dépendent, au cours de la période de thermosensibilité, des fractions du développement passées en dessous ou au dessus de la TRT.
La différenciation et la croissance des gonades dépend de la température
Les modifications morphologiques majeures qui apparaissent pendant la période de différenciation des gonades sont assez semblables chez toutes les espèces de reptiles.
Comme chez les autres vertébrés, la crête génitale se développe par épaississement de l'épithélium coelomique, sur la face ventro-médiane du mésonephros. Les cellules germinales primordiales sont dispersées dans cet épithélium (épithélium germinatif). Au cours du développement de ces crêtes génitales, des cellules épithéliales s'ajoutent au mésenchyme initial au centre de la gonade (medulla). Certaines de ces cellules (provenant de corpuscules de Malpighi ou de l'épithélium coelomique) formeront les cordons du rete. D'autres (provenant de l'épithélium germinatif lui-même) donnent des cordons épithéliaux qui pénètrent dans le mésenchyme sous-jacent. Ces cordons médullaires (ou cordons sexuels) s'entourent rapidement d'une membrane basale. Les cordons sexuels et ceux du rete entrent en contact dans la partie dorsale de la gonade. Cette structure de la gonade est observable dans les deux sexes avant la période thermosensible et à ce stade, la gonade est considérée comme indifférenciée. La différenciation sexuelle gonadique se fera pendant la période thermosensible.
|
|
|
Différenciation gonadique chez les reptiles présentant une détermination du sexe par la température. a, albuginée; c, cortex; cB, capsule de Bowman du corpuscule de Malpighi; cg, cellule germinale; cL, cellule de Leydig; cm, cordon médullaire; cr, cordon du rete; ec, épithélium coelomique; ecr, ébauche de cordon du rete; ecs, ébauche de cordon séminifère; eg, épithélium germinatif; fp, follicule primordial; l, lacune; m, medulla; md, mésentère dorsal; mm, mésenchyme mésonéphritique; o, ovocyte; tm, tube mésonéphritique; vs, vaisseau sanguin |
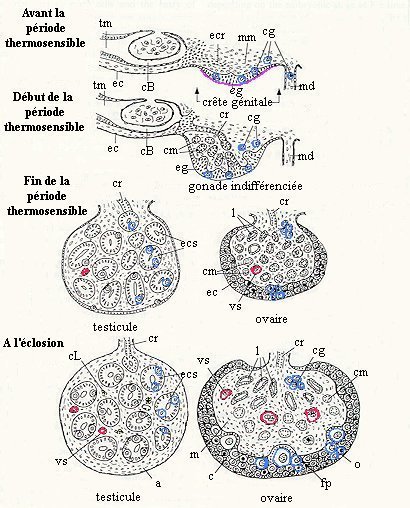
La différenciation dans le sens mâle commence par un amincissement de l'épithélium germinatif. Les cellules germinales le quittent et migrent jusqu'à s'insérer entre les cellules épithéliales des cordons médullaires. Dans ces cordons un nombre de cellules épithéliales de plus en plus important acquiert les caractéristiques "Sertoliennes". Les cordons médullaires forment ainsi les ébauches des cordons séminifères. Pendant la période thermosensible, sous l'effet du développement de ces cordons séminifères dans la medulla, la croissance des testicules est régulière. Les cellules de Leydig ne se différencient, dans le tissu interstitiel, qu'après la période thermosensible.
Dans les ovaires en cours de différenciation, l'épithelium germinatif s'épaissit sous l'effet de la prolifération, in situ, de cellules épithéliales et germinales qui donneront naissance aux follicules. Ainsi, dans l'ovaire, c'est essentiellement le cortex qui se développe. Durant la période thermosensible, l'ovaire grossit moins que le testicule, sa medulla étant plus réduite. A la fin de la période thermosensible, certaines cellules germinales sont entrées en méiose et à l'éclosion, on observe généralement quelques follicules primordiaux avec leurs ovocytes en croissance, dans la zone interne du cortex ovarien.
Les stéroïdes interviennent dans la différenciation gonadique
Au début des années 70, lorsqu'on découvrit le déterminisme du sexe par la température chez les tortues, il était bien établi que des traitements d'embryons ou de larves par des oestrogènes de synthèse pouvaient féminiser des mâles génétiques, chez les oiseaux, les reptiles (lézards), les amphibiens et les poissons. Chez les marsupiaux, on avait obtenu le développement d'ovotestes par un traitement précoce aux oestrogènes de jeunes mâles encore dans la poche; cependant , chez les euthériens, le traitement de femelles gravides par les oestrogènes ou les androgènes ne modifiait pas la différenciation sexuelle des gonades des foetus.
C'est dans ce contexte que fut étudié l'effet de l'injection d'androgène (propionate de testostérone) ou d'oestrogène (benzoate d'oestradiol) dans des oeufs de tortues (Emys orbicularis, Testudo gracea).
A une température déterminant le sexe femelle, la testostérone n'inverse pas le phénotype sexuel de la gonade. Mais à une température déterminant le sexe mâle, elle induit la formation d'un cortex de type ovarien, à la surface du testicule (effet féminisant dit "paradoxal").
A une température déterminant le sexe mâle, l'oestradiol induit différents degrés de féminisation des testicules, allant de l'ovotestis à l'ovaire, ces divers degrés dépendant du stade embryonnaire au moment de l'injection et de la dose injectée.
Les grandes similitudes structurales entre ovaires obtenus sous l'effet de la température ou induits par l'oestradiol sont frappantes. De même les ressemblances entre ovotestes obtenus par injection hormonale et gonades intersexués obtenues aux températures pivots indiquent une étroite relation entre la structure de la gonade et le taux d'oestrogène.
Des expériences sur les tortues (notamment Emys orbicularis) utilisant des oestrogènes, des antioestrogènes ou des inhibiteurs de l'aromatase (l'aromatase permet la transformation des androgènes en oestrogènes) ont donnés des résultats qui prouvent l'intervention précoce des oestrogènes dans la différenciation de l'ovaire.
|
Température d'incubation |
Traitement |
Structure de la gonade |
|
25°C |
aucun = témoin |
testicule |
|
idem |
benzoate d'oestradiol |
ovaire |
|
idem |
antioestrogène (tamoxifène) |
ovotestis |
|
idem |
inhibiteur de l'aromatase (fadrozole ou létrozole) |
testicule |
|
30°C |
aucun = témoin |
ovaire |
|
idem |
benzoate d'oestradiol |
ovaire |
|
idem |
antioestrogène (tamoxifène) |
ovotestis |
|
idem |
inhibiteur de l'aromatase (fadrozole ou létrozole) |
ovotestis ou testicule |
Les cas où la gonade présente une intersexualité (ovotestis) peuvent se rencontrer dans la nature: des testicules fertiles présentent alors à leurs suface des ovocytes immatures. L'obtention de ce genre de gonades, naturellement, à la température pivot, ou sous l'effet de traitements est expliquée par une légère croissance du taux d'oestrogènes qui est alors suffisant pour induire la formation d'un cortex de type ovarien mais insuffisant pour inhiber le développement de cordons testiculaires. En général, après l'éclosion les taux d'oestrogènes sont insuffisants pour maintenir le cortex ovarien mais parfois quelques cellules germinales peuvent exister encore dans les vestiges du cortex de l'ovotestis.
L'aromatase est le complexe enzymatique qui convertit les androgènes en oestrogènes (l'androstènedione en oestrone et la testostérone en oestradiol). Dans les gonades d'embryons de tortue incubés à 25°C (TM), elle présente une activité très faible durant la période thermosensible et jusqu'à l'éclosion. Alors que dans les gonades d'embryons incubés à 30°C (TF) l'activité de l'aromatase est également très faible au début de la période thermosensible mais augmente de façon exponentielle durant cette période; elle présente une légère diminution au moment de l'éclosion. L'augmentation de l'activité de l'aromatase peut donc être mise en relation avec la féminisation de la gonade.
Dans le cas des ovotestes, et de l'effet féminisant paradoxal de la testostérone, il est possible que l'administration de testostérone exogène en quantité considérable par rapport au niveau normal, active sensiblement, par un mécanisme encore inconnu, la synthèse d'aromatase gonadique.
Une autre enzyme, la 3bêtaHSD (3bêta-hydroxystéroide-déshydrogénase-5ène-4ène-isomérase) qui permet la synthèse de progestérone et d'androgènes à partir du cholestérol, pourrait être impliquée dans le déterminisme thermique du sexe.
On a montré que la 3bêta HSD est présente, avant la période thermosensible, dans les cordons épithéliaux médullaires de la gonade indifférenciée. Cependant, durant la période thermosensible, son activité augmente dans les testicules (cordons testiculaires) en différenciation à une température déterminant le sexe mâle (25°C pour E.orbicularis) alors qu'elle diminue dans les ovaires (medulla) en différenciation à une température déterminant le sexe femelle (30°C pour E.orbicularis). Ainsi, dans les gonades, la synthèse de cette enzyme de la stéroidogénèse est influencée par la température alors qu'elle ne l'est pas dans la corticosurrénale.
Ainsi chez cette tortue, et d'autres espèces présentant un déterminisme thermique du sexe, deux enzymes clé de la stéroidogénèse présentent une activité qui dépend de la température, durant la période de sensibilité à ce facteur. Si l'implication de la 3bêtaHSD dans le déterminisme thermique du sexe demeure inconnue, en revanche, celle de l'aromatase dans la féminisation de la gonade, via les oestrogènes, est démontrée.
Contrôle génétique du sexe et influence de la température
La recherche, chez les espèces présentant un déterminisme thermique du sexe, de gènes homologues à ceux impliqués dans le déterminisme du sexe chez les mammifères a montré de nombreuses ressemblances.
Deux gènes; WT1 et SF1, sont impliqués dans la formation des crêtes génitales (et la stéroïdogénèse pour SF1) chez les mammifères. Des homologues de ces deux gènes ont été trouvés chez les reptiles.
Chez les mammifères, la différenciation testiculaire est gouvernée par le gène SRY. Plusieurs autres gènes sont associés à la différenciation des cellules de Sertoli; en particulier les gènes SOX9 (de la même famille que SRY) et DMRT1 ainsi que le gène de l'AMH (hormone anti-Müllérienne).
On a trouvé chez les reptiles plus de 10 gènes de la famille SOX, mais pas le gène SRY qui semble spécifique des mammifères. Récemment deux transcrits du gène SOX9 ont été identifiés chez une tortue, l'un de 3.9kB et l'autre de 2.2kB (le deuxième correspondant à une coupure du premier). Ces deux transcrits se rencontrent dans l'ensemble mésonéphros/gonade et dans le cerveau à toutes les périodes du développement mais durant la période thermosensible les proportions de ces deux transcrits varient selon la température ambiante. Enfin l'expression de SOX9 a été examinée par hybridation in situ (sonde ARN anti-sens) et par immunohistochimie (anticorps anti-protéine SOX9); ces examens ont montré qu'avant et au début de la période thermosensible ce gène s'exprime dans les gonades aux deux températures d'incubation, mais qu'à la fin de cette période il est exprimé seulement dans les testicules et pas dans les ovaires, alors qu'il s'exprime dans le mésonéphros à toutes ces périodes.
DMRT1 pourrait jouer un rôle clé dans la différenciation testiculaire des reptiles présentant un déterminisme thermique du sexe. En effet, s'il est exprimé dans les gonades indifférenciées (avant la période thermosensible), à température féminisante comme à température masculinisante, il ne l'est plus dans les ovaires en cours de différenciation (pendant la période thermosensible) alors que son expression augmente dans les testicules pendant la même période.
Le gène de l'AMH a été également trouvé, il présente de nombreuses similitudes avec celui des mammifères, mais contrairement à ces derniers, chez l'alligator, son expression dans les testicules précède celle de SOX9. Récemment, l'homologue du gène DAX1 des mammifères a été également mis en évidence chez l'alligator.
Ainsi, il apparaît, qu'à l'exception du gène SRY, tous les gènes intervenant dans la détermination du sexe chez les mamifères ont leurs homologues chez les reptiles dont le sexe est déterminé par la température.
Mais plusieurs questions restent posées:
- ces gènes interviennent-ils ici directement ou indirectement?
- sont-ils contrôlés par les oestrogènes?
- comment la température intervient-elle dans ce processus?
Interactions entre ces différents gènes
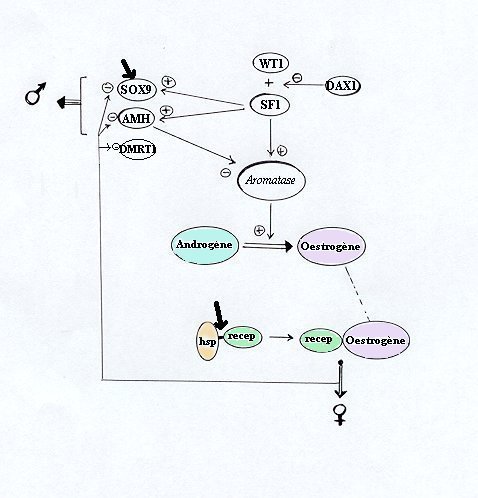
Chez les mammifères, SF1 agit comme activateur de la transcription de certains gènes codant pour les enzymes de la stéroïdogénèse, comme l'aromatase. Ce gène SF1 est également impliqué dans l'activation des gènes SOX9 et AMH, tous deux spécifiques de la différenciation testiculaire, avec une amplification de cette transcription quand WT1 interagit avec SF1.
D'un autre côté on sait que l'AMH inhibe l'expression de l'aromatase.
On peut alors supposer que, chez les reptiles dont le sexe dépend de la température, SF1 peut agir comme un facteur masculinisant par l'intermédiaire de SOX9 et AMH ou comme un facteur féminisant par activation de l'aromatase.
DAX1 qui, chez les mammifères, empêche la synergie entre WT1 et SF1, réprimant ainsi la transcription de SOX9 et AMH, peut être considéré comme un facteur féminisant. On ne sait pas si il a le même rôle chez les reptiles ni s'il est impliqué dans le contrôle de l'aromatase par SF1.
Des interactions de DMRT1, facteur masculinisant, sont possibles avec l'un ou plusieurs de ces facteurs.
Le modèle proposé ici suppose que la transcription des gènes masculinisant et féminisant est sous le contrôle des oestrogènes, les gènes masculinisants étant inhibés par ces oestrogènes et les gènes féminisants étant activés. Cependant la cible précise de la température dans ce processus n'est pas réellement connue.
En effet, la température peut inhiber ou activer les gènes de facteurs féminisant ou masculinisant, par exemple en agissant sur SOX9, ou sur les protéines de choc thermique (heat-shock protéin, hsp) impliquées dans la liaison des oestrogènes à leur(s) récepteur(s). Elle peut être impliquée dans la dissociation de la protéine de choc thermique du complexe oestrogène-récepteur qui sera alors activé.
|
|
|
Interactions possibles des homologues des facteurs de détermination du sexe des mammifères avec le gène de l'aromatase chez les reptiles présentant un déterminisme thermique du sexe et cibles possibles de la température (flèches épaisses). hsp, protéine de choc thermique ; recep, récepteur des oestrogènes |