
Organisation fonctionnelle du cortex visuel primaire (aire V1)
Localisation et structure histologique
Le cortex visuel primaire (ou cortex strié ou aire 17 de la classification de Brodmann, ou aire V1) se situe aux pôles postérieurs des lobes occipitaux. Il reçoit diverses informations concernant le monde visuel. Ces informations lui parviennent par des canaux séparés, déjà décomposées en un certain nombre de caractérisques.
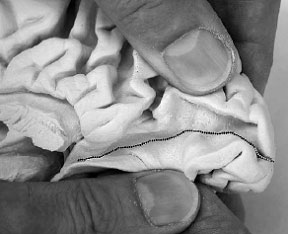
Anatomiquement le cortex visuel primaire se situe sur les berges de la fissure calcarine avec laquelle il s'enfonce dans la profondeur du lobe occipital.
  |
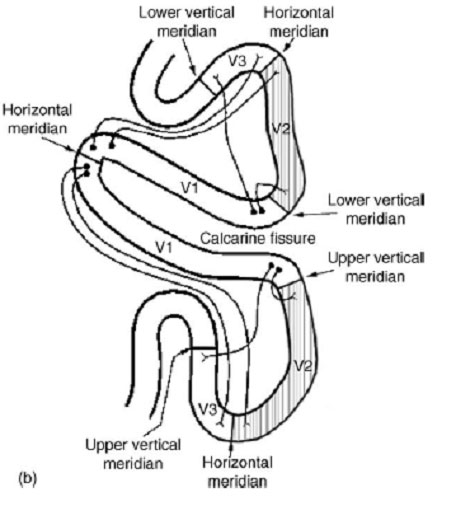 |
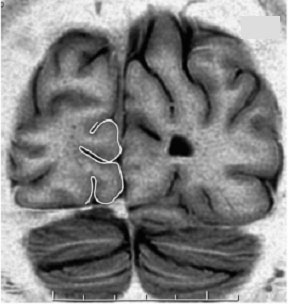
| Situation macroscopique de l'aire V1 au niveau de la fissure calcarine (surlignée en noir) et localisation des aires V1 V2 et V3 (surlignées en blanc) sur une image IRM , en coupe coronale. | Position des aires V1, V2 et V3 sur un croquis de la coupe du cortex (même position que sur limage IRM) |
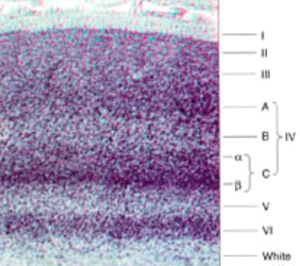
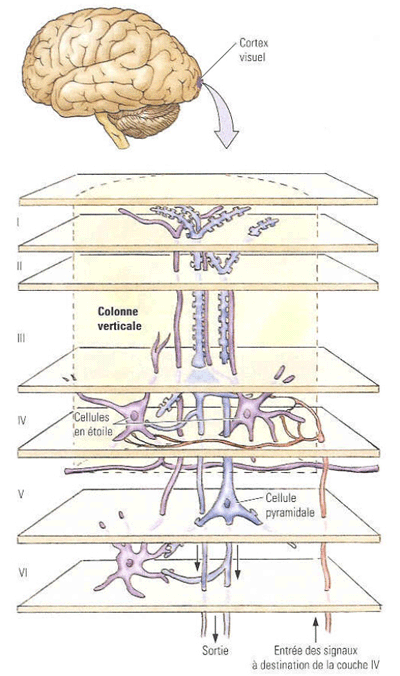
Microscopiquement, il est formé , comme tout néocortex, de 6 couchescellulaires, et est aisément reconnaissable à l'oeil nu par la présence en son milieu d'une strie grise (strie de Gennari) correspondnat à une couche granulaire externe (couche IV) particulièrement développée.
 |
 |
|
Figure 1. Coupe histologique colorée du néocortex dans la region V1 La plupart des terminaisons des neurones provenant du corps genouillé latéral (CGL) se font au niveau de la couche IV de l'aire V1 (en majorité dans la couche IVc) où elles réalisent une synapse avec les petites cellules étoilées, elles-mêmes en contact avec les couches voisines.
|
Figure 2. Représentation des différentes types cellulaires des couches de l'aire V1 Dans la couche IVc, la plupart des cellules étoilées ont des champs récepteurs du type ON et du type OFF
|
Organisation fonctionnelle de l'aire V1
De même que Stephen Kuffler avait exploré les propriétés réactionnelles des cellules ganglionnaires, David Hubel et Torsten Wiesel ont examiné, à l'aide d'enregistrements par micro-électrodes, les propriétés des neurones de structures visuelles plus centrales, comme l'aire visuelle primaire, au niveau du cortex. Ce sont les travaux de Hubel et Wiesel qui sont à la base de nos connaissances sur l'organisation focntionnelle du cortex strié (aire V1).
La sensibilité à l'orientation
Les réponses des neurones du corps genouillé latéral sont similaires à celles des cellules ganglionnaires de la rétine. On retrouve une organisation du champ récepteur en centre et pourtour et une sélectivité aux variations de luminance notamment (augmentations ou diminutions). Par contre, dans le cortex strié, les étroits faisceaux lumineux qui stimulaient si efficacement les neurones de la rétine et du CGL deviennent en grande partie inefficaces.
Il a été montré, chez le chat et le singe, que certains neurones corticaux répondent fortement à des bords ou des barres de contraste, à condition qu'ils soient présentés dans le champ récepteur selon une orientation particulière. Chaque neurone présente une "orientation préférentielle" pour laquelle sa décharge de potentiels d'action est maximale. Toutes les orientations d'un bord ou d'une barre sont représentées de façon à peu près égale dans le cortex. Le codage d'une orientation particulière d'un élément d'une scène visuelle se fait par l'activité d'une population spécifique de neurones de V1.
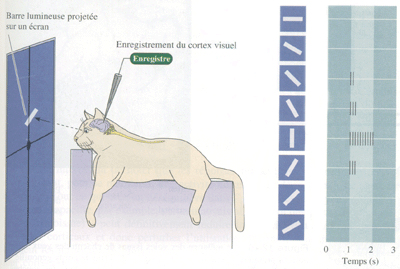 |
Figure 3. Enregistrement des réponses de neurones corticaux sensibles à l'orientation (Neurosciences, Purves) On projette devant les yeux de l''animal une barre lumineuse d'orientation variable, durant une seconde (entre 1ère et 2 ème seconde), et on enregistre les réponses électriques (portentiels d'action) de cellules corticales.
|
Deux types cellulaires ayant une sensibilité à l'orientation ont été décrits.
-
Des cellules "simples" (principalement dans la couche IV) ayant un champ recepteur fait d'une zone centrale excitatrice et d'une zone périphérique inhibitrice, mais dont la zone centrale (au lieu de circulaire comme pour les cellules rétiniennes ou du CGL) présente une forme allongée. Le stimulus pertinent est une barre lumineuse d'une orientation donnée (l'orientation dans le sens de l'allongement du champ récepteur entraîne le maximum de réponse), seulement s'il occupe une position précise dans le champ récepteur (à cause de la zone périphérique inhibitrice).
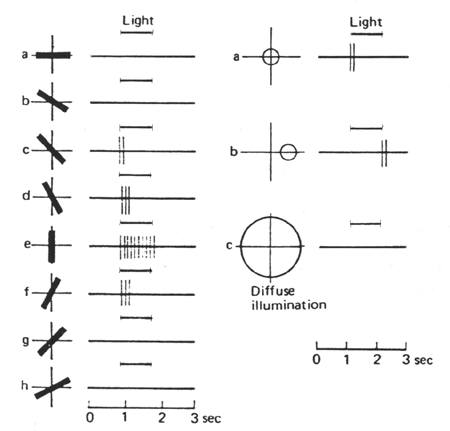
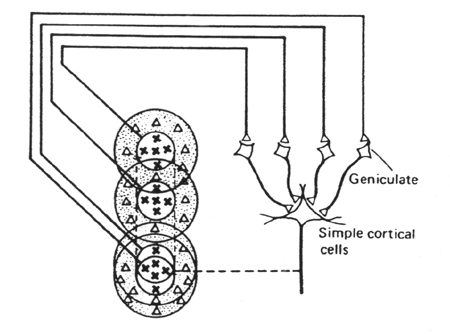
Figure 4. Etude du champ récepteur d'une cellule simple
L'illumination du champ récepteur d'une cellule simple par une barre lumineuse n 'entraîne une activation de la cellule corticale que lorsque l'orientation de la barre se rapproche de l'orientation spécifique de la cellule (ici la verticale).
Figure 5. Interprétation intégrative du champ récepteur d'une cellule simple
Schéma de Hubel et Wiesel expliquant les propriétés du champ récepteur d'une cellule simple. Cette cellule reçoit des connexions excitatrices de 4 ou davantage de cellules du CGL, ayant des organisations similaires de leur centre et pourtour, et disposés en ligne verticale au niveau de la rétine.
Si une barre lumineuse éclaire à la fois plusieurs centres ON (marqués de croix) de cellules gangglionnaires, il y aura excitation de plusieurs cellules du CGL et forte excitation de la cellule simple (stimulation de son centre ON de plus grande taille et de forme allongée). -
Des cellules "complexes" (principalement hors de la couche IV) possèdent également un axe d'orientation spécifique, mais elles ont un champ récepteur plus vaste et celui-ci ne possède pas de zone inhibitrice périphérique. Elles présentent un mélange de réponse ON et OFF dans toute l'étendue de leur champ récepteur, comme si elles recevaient leurs afférences de plusieurs cellules simples. Elles sont également sensibles à l'orientation d'un stimulus, quelque soit sa position dans le champ visuel. En outre, ces cellules complexes répondent au mouvement d'un stimulus orienté si ce mouvement est perpendiculaire à l'axe de son orientation.
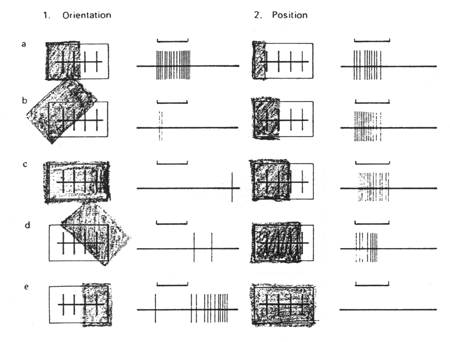
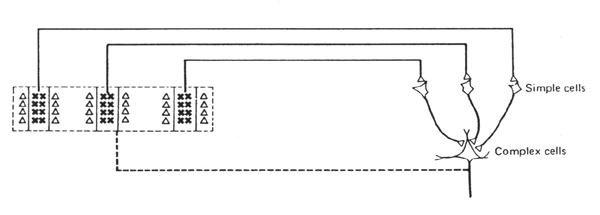
Figure 6. Etude du champ récepteur d'une cellule complexe
Une cellule complexe répond spécifiquement à l'orientation du bord du stimulus mais non à sa position dans le champ visuel; une orientation de ce bord autre que vertical est pratiquement inefficace. Une source lumineuse (zone crayonnées) ayant ici son bord vertical à droite est excitatrice, une source ayant son bord vertical à gauche est inhibitrice. Une illumination totale du champ récepteur entraîne une absence de réponse.Figure 7. Interprétation intégrative du champ récepteur d'une cellule complexe
Schéma de Hubel et Wiesel expliquant les propriétés du champ récepteur d'une cellule complexe du type de celle ci-contre. Cette cellule reçoit des connexions excitatrices de plusieurs cellules simples qui présentent un axe d'orientation spécifique vertical, avec une zone centrale excitatrice ( marquée de croix) et deux zones latérales inhibitrices, et dont les champs récepteurs sont disposées en ligne horizontale sur la rétine.
Des analyses ultérieures ont également mis en évidence des neurones corticaux sensibles à la longueur d'une barre lumineuse se déplaçant dans le champ récepteur, neurones dont la fréquence de décharge diminue quand cette barre dépasse une certaine longueur.
Les colonnes de dominance oculaire
La démonstration de la structure columnaire du cortex a été faite grâce à l'injection au niveau d'un oeil chez le singe) d'un acide aminé radioactif et par analyse autoradiographique de coupes hitologiques du cortex visuel. Hubel et Wiesel ont ainsi constaté que la substance radioactive se fixait dans la couche IV (au niveau des terminaisons des fiobres en provenance du CGL) selon une topographie particulière, réalisant une séries de stries alternantes sombres (non marquées par la substance radioactive) et claires (marquées) de 0,4 à O,5 mm de largeur.
 |
| Figure 8. Injection de l'acide aminé radioactif dans l'un des deux globes oculaires d'un singe (Hubel et Wiesel) |
 |
 |
| Figure 9. Autoradiographie d'une coupe histologique du cortex visuel primaire | Figure 10. Reconstitution tridimensionnelle d'une tranche de cortex visuel primaire (couche IV de l'aire V1) |
Cette organisation du cortex primaire en colonnes de dominance oculaire témoigne de l'arrivée à chaque aire V1, d'informations en provenance des deux yeux.
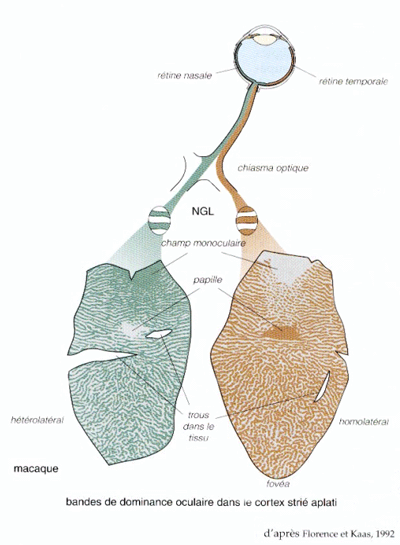 |
| Figure 11. Bandes de dominance oculaire dans les deux aires V1 après injection de traceurs radioactifs dans un seul des globes oculaires (La vision, Rodieck) |
La vision en relief
Chaque corps genouillé latéral reçoit des afférences des deux yeux et celles-ci, comme au niveau de la couche IV du cortex, sont séparées. Ces neurones sont monoculaires (ils ne répondent à la stimulation que d'un seul oeil, droit OU gauche). Les neurones de la couche IV émettent leurs axones vers d'autres couches corticales. C'est à cette étape que les messages des deux yeux convergent sur les mêmes neurones individuels.
Au dela de la couche IV, les neurones du cortex visuel primaire sont tous binoculaires, les signaux des deux yeux se combinent au niveau d'une même cellule. Ainsi la plupart des neurones corticaux ont des champs récepteurs binoculaires et ces champs sont presque identiques (ils ont la même taille, la même forme, la même orientation préférentielle et à peu près la même position dans l'espace visuel de chaque oeil.
La réunion, dans le cortex strié, des afférences venant des deux yeux est la base de la stéréopsie, c'est à dire de la sensation de profondeur ou de relief qui se manifeste quand on regarde des objets proches avec les deux yeux et non un seul. Etant donné que les deux yeux voient le monde sous des angles légèrement différents, les objets qui se situent en avant ou en arrière du plan de fixation se projettent sur des points non homologues de chacune des rétines.
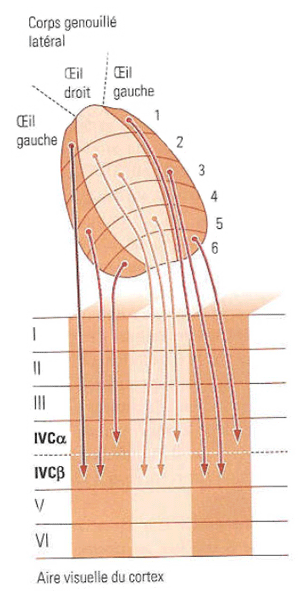 |
Figure 12. Projection des neurones du corps genouillé latéral vers les couches du cortex visuel primaire Les cellules P (parvocellulaires) et M (magnocellulaires) se terminent dans deux bandes distinctes de la région IV, les cellules P dans la couche IVC-alpha et les cellules M dans la couche IVC-beta. Le cortex visuel primaire gauche (par exemple) reçoit l'ensemble des neurones correspondant à l'hémichamp droit, c'est à dire les neurones issus de la rétine temporale gauche et de la rétine nasale droite.
|
Ainsi sur une petite distance en avant et en arrière du point de fixation où la disparité entre les vues du monde par les deux yeux demeure modeste, on ne voit qu'une seule image et la différence perçue entre les deux yeux est interprétée comme une profondeur.
Les bases physiologiques de la stéréopsie ne sont pas encore connues, mais certains neurones existant dans le cortex strié et d'autres aires visuelles sont de bon candidats. Contrairement aux cellules binoculaires du cortex visuel primaire, dont les champs monoculaires concernent la même région de l'espace, ces neurones ont des champs monoculaires légèrement décalés et ils subissent une excitation maximale de la part de stimulus qui "tombent" sur des endroits non homologues des deux rétines. Certains (les cellules "loin") déchargent pour des stimuli placés au delà du point de fixation, d'autres (les cellules "près") répondent à des stimuli situés en deça du plan de fixation.
Une organisation "modulaire"
Les cellules d'orientation sont organisées en colonnes (une colonne regroupe des cellules sensibles à la même orientation) d'environ 50 micomètres d'épaisseur. Ce sont donc des colonnes plus étroites que les colonnes de dominance oculaire et elles sont disposées perpendiculairement à ces dernières.
Ces colonnes d'orientation sont, de plus, disposées selon un ordre très précis: l'axe auquel les cellules sont sensibles suit progressivement, de colonne en colonne une succession géométrique de 10° environ.
Enfin une troisième organisation tridimensionnelle existe dans le cortex visuel primaire. On trouve une disposition en cylindres (formant des taches nettes lors de coupes du cortex à travers ses couches II et III notamment) de cellules sensibles à la couleur (regroupées dans les blobs), alors que les cellules sensibles à la forme sont entre ces "blobs", dans les zones inter-taches. Voir l'organisation des aires visuelles.
Finalement l'image d'un point du champ visuel est traité par une portion de 2mm x 2mm de cortex visuel de l'aire V1, renfermant:
-
2 sets de colonnes de dominance oculaire (couche IV)
-
2 sets de colonnes d'orientation de 0° à 180° (couche III)
-
16 blobs sensibles à la couleur (couche III)
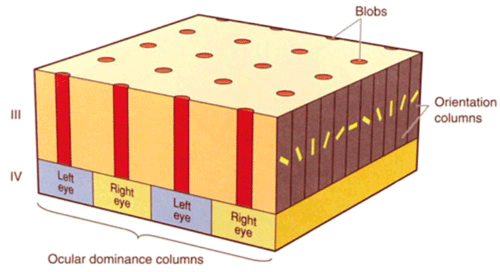 |
| Figure 13. Un module cortical |
C'est ce que l'on nomme un module cortical (ou hyper-colonne) et c'est un volume qui paraît nécessaire et suffisant pour analyser un point de l'espace visuel.