
Traitement rétinien de la couleur
La rétine capte le spectre lumineux provenant de l'objet au niveau de sa couche de cellules photoréceptrices. Celles-ci élaborent des signaux qui ne sont pas des messages nerveux mais s'apparentent à des potentiels de récepteur. C'est au sortir de cette rétine, que le message, codé en fréquence de potentiels d'action, sera transmis des cellules ganglionnaires aux structures cérébrales (corps génouillé latéraux et cortex visuel).
Au cours de ce passage à travers les différents types cellulaires de la rétine, il s'opère un traitement de l'information qui constitue un premier codage de l'image.
L'étage des photorécepteurs
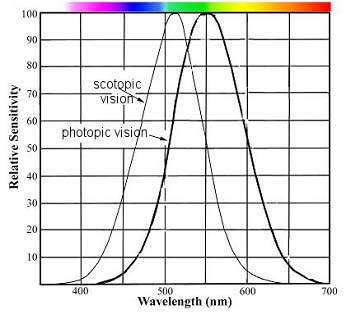
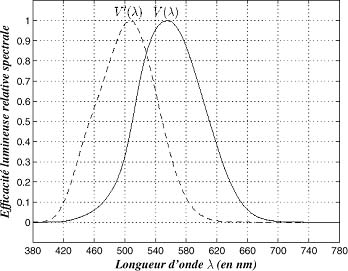
La réponse de l'oeil à une lumière monochromatique a été mesurée en fonction de la longueur d'onde. La CIE (Commission Internationale de l'Eclairage) a admis une réposne type correspondant à un observateur moyen. Elle est représenté ci-dessous pur deux types de vision :
- diurne ou photopique (où le maximum de sensibilité se trouve à 555 nm (couleur verte)
- pré-nocturne ou scotopique (avec un maximum de sensibilité décalé vers le bleu)
 |
 |
|
Courbe de sensibilité de l'oeil selon la longueur d'onde. Cette fonction appelée aussi V(lamba) peut être normalisée à 1 en son maximum, comme présenté dans la figure de droite |
|
Les deux grands types de photorécepteurs de la rétine, les batonnets et les cônes, présentent des caractéristiques fort distinctes.
Les bâtonnets qui sont très nombreux et plus sensibles à la lumière que les cônes, sont responsables de la vision scotopique (ils sont saturés en vision photopique), leur temps d’adaptation aux changements de conditions est par contre beaucoup plus long que celui des cônes. Avec un seul type de pigment (la rhodopsine), ils ne peuvent distinguer les couleurs et sont sensible uniquement à la luminance. De par leur répartition hors de la fovéa, ils sont responsables de notre vision périphérique.
Les cônes, beaucoup moins nombreux et essentiellement répartis dans la fovéa, sont responsables de la vision haute résolution. Ils sont moins sensibles à la lumière que les bâtonnets, ils fonctionnent en vision photopique, mais peuvent s’adapter très rapidement à des changements d’intensité.
On distingue ainsi 3 types de cônes : les cônes S sensibles à des longueurs d’onde courtes (short), les cônes M sensibles à des longueurs d’onde moyennes (medium) et les cônes L sensibles à des longueurs d’onde longues (long). C’est là l’origine de l’aspect trichromatique de la vision des couleurs.
Les cônes L sont sensibles au jaune-vert à rouge, les cônes M au vert et les cônes S au bleu.
Les cônes S sont les moins nombreux : 64 % L, 32 % M et seulement 2% S, bleus.
Les pigments (des protéines) des cônes sont appelés cyanolabe, chlorolabe et erythrolabe dans les cônes S, M et L respectivement.
Les cônes S sont particuliers. Ils sont plus sensibles à la lumière, peu nombreux dans la fovea: ils sont responsables de certains phénomènes d’aberration chromatique.
 |
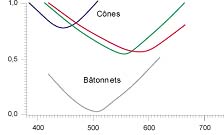 |
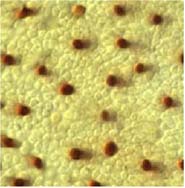
| Répartitionn des cônes (sombre) et des batonnets (clairs) dans la rétine, en dehors de la fovea. | Seuil de sensibilité en fonction de la longueur d'onde des 4 types de photorécepteurs (Perception humaine de la couleur, J-M Frigerio). |
Les voies M,P,K et la vision des couleurs
A l'intérieur de la rétine, les différents cônes répondent à une stimulation lumineuse en donnant naissance à des potentiels de récepteurs d'amplitude variable et correspondant à leur degré de sensibilité pour la longueur d'onde de ce rayon électromagnétique (en cas de source lumineuse mixte les potentiels de récepteurs correspondant à la réponse vis à vis des deux (par exemple) fréquences s'additionnent. Ainsi il y a une perte de l'information correspondant à la longueur d'onde elle-même.
Ces potentiels ne sont pas des messages, seuls les influx nerveux codés en fréquence de potentiels d'action et qui sont produits par les cellules ganglionnaires de la rétine sont de véritables messages qui seront propagés vers les aires visuelles corticales primaires, après relai dans les corps genouillés latéraux (CGL).
On admet actuellement, en fonction des maxima de réponses enregistrées dans les différents types cellulaires du CGL, qu'il existe dans la rétine un fonctionnement en réseau (impliquant photorécepteurs, cellules bipolaires et cellules ganglionnaires) assurant une sommation des réponses de différents cônes, selon le schéma suivant.
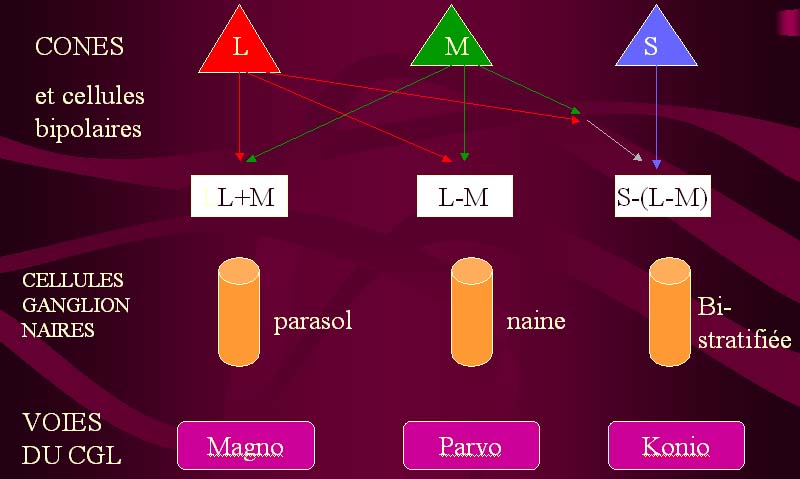 |
| Schématisation de l'hypothèse actuelle sur le traitement de l'information colorée dans les différents étages de la rétine. |
Ce système, basé sur la comparaison d'intensité de réponse pour des couples antagonistes de couleurs (couple rouge-vert et couple belu-jaune) a pour effet principal d'augmenter le pouvoir discriminateur de l'oeil dans la vision colorée . Pour deux fréquence proches, la différence de réponse au sortir des cellules ganglionnaires des voies M, P et K est plus importante qu'au sortir des cônes.
Ce système permet de comprendre pourquoi une teinte semble à nos yeux conserver sa couleur (principe d'univariance) malgré des différentces permanentes de son éclairage, puisque la vision des couleurs est ici basée sur un système de comparaison de réponses et non sur un système de réponse absolue à une fréquence donnée.