
Genétique de la vision colorée
Bases génétiques de la vision des couleurs
Ecrit par Françoise Jauzein
D'après Dossier hors série Pour La Science "La couleur", avril 2000
Bref historique
La description de la vision des couleurs a commencé il y a plusieurs siècles. Une première contribution importante fut apportée par Isaac Newton qui découvrit le spectre de la lumière. Il observa également que l'oeil humain ne distingue pas des couleurs formées par combinaisons de rayonnements de couleurs pures: ainsi, une couleur formée d'un mélange de rouge et de vert apparaîtra aussi "jaune" qu'une couleur jaune "pure". Des couleurs différentes perçues de la même façon sont dites métamères.
A la fin du XVIIIème siècle, on découvrit que la vision des couleurs est trichromatique: on obtient la plupart des sensations colorées en mélangeant les trois couleurs primaires. De nombreuses lumières pures (monochromatiques) peuvent servir de lumières primaires mais tous les jeux de trois couleurs primaires comportent une lumière de grande longeur d'onde, une de moyenne longueur d'onde et une de courte longueur d'onde : rouge/vert/bleu (pour les écrans) ou cyan/jaune/magenta (dans l'imprimerie).
En 1802 le médecin et physicien anglais Thomas Young imagina que cette trichromie était en relation avec la façon dont nous percevons les couleurs: "comme il est impossible d'imaginer que chaque point photosensible de la rétine contienne une infinité de particules susceptibles de vibrer à l'unisson avec tous les rayonnements, il faut postuler l'existence de seulement trois types de particules, sensibles aux trois couleurs primaires". La communauté scientifique mit du temps à l'admettre!
Aujourd'hui, on sait que les trois classes de cellules photosensibles de la rétine, les cônes, ont des sensibilités différentes à la lumière, même si leur domaine de sensibilité se recouvrent (les récepteurs sensibles au vert absorbent également la lumière orangé). Ce qui caractérise une teinte n'est donc pas l'intensité de réponse d'un type de cône, mais l'ensemble des niveaux de réponse des trois types de cônes. C'est ainsi qu'une lumière composée d'une certaine intensité de deux fréquences différentes pourra exciter les trois types de cônes de la même façon qu'une seule fréquence (lumière pure).
Paralèllement aux travaux de Young , John Dalton, un chimiste, rapporta , dès 1794, qu'il ne voyait pas les couleurs comme tout le monde : "les parties d'image que les autres voient rouges m'apparaissent comme des ombres ou des absences de lumière, l'orangé , le jaune et le vert sont pour moi comme plusieurs intensités de jaune".
On appelle daltonisme la déficience de l'aptitude à distinguer les couleurs de la partie du spectre qui s'étend du rouge au vert (8% environ des hommes et 1% environ des femmes). Certains individus distinguent mal les couleurs de la partie bleue du spectre, ils sont très rares.
Au milieu du XIXème siècle, le physicien anlais James Clerck Maxwell identifia deux types de daltonisme en présentant plusieurs couleurs à des sujets et en examinantsystématiquement les couleurs qu'ils ne distinguaient pas. En appliquant la théorie des trois types de récépteurs de Young, il mit en évidence deux types de déficience:
- les personnes confondant les couleurs excitant autant les récepteurs sensibles au rouge et ceux sensibles au bleu
- les personnes confondant les couleurs excitant autant les récepteurs sensibles au vet et ceux sensibles au bleu
Il supposa que les personnes normales distinguaient ces couleurs grâce à des différences d'excitation des récepteurs sensibles au vert (dans le premier cas) ou au rouge (dans le second cas). Il en déduisit que les personnes du premier groupe ne possédaient pas les récepteurs sensibles au vert et que celles du second groupe ne possédaient pas les récepteurs sensibles au rouge. Ce sont respectivement des dyschromates vert moins (daltonisme deutéranope) et des dyschromates rouge moins (daltonisme protanope).
A la fin du XIXème siècle fut mit au point un appareil (par le matématicien et physicien anglais John William Strutt, dit Lord Rayleigh) permettant de tester la vision des couleurs dans la zone rouge-vert: l'anomaloscope.
Son principe repose sur la comparaison sur un écran d'une lumière "pure" jaune orangée avec une lumière composée d'une addition d'une certaine intensité de rouge et de vert. Les sujets normaux sont sensibles au rouge et au vert et ajustent ces deux intensités (plus de rouge que de vert) pour obtenir un jaune comparable à la lumière pure. Les sujets dyschromates "rouge moins" n'ajustent cette lumière qu'en modifiant l'intensité du vert, et les dyschromates "vert moins" qu'en modifiant le rouge.
Rayleigh, avec cet appareil, découvrit un autre type d'anomalie. Certaines personnes jouaient bien sur le rouge et le vert pour équilibrer le jaune orangé, mais le faisaient dans des proportions anormales. Il en conclut que les pectres d'aborption de leurs récepteurs rouge ou vert étaient anomaux (maximum de sensibilité décalé). Ce sont des trichromates vert anormaux ou rouge anormaux.
En 1960, des microspectrophotomètres déterminant l'absorption d'une seule cellule photoréceptrice permirent de comparer les intensités d'absorption des différentes longeurs d'onde par les cônes et de confirmer l'hypothèse des trois types de cônes émise par Young.
courbes de sensibilité des pigments
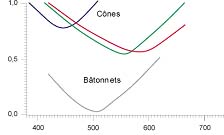
Trois types de pigments dans les cônes: les cônes L sont sensibles aux grandes longeurs d'ondes (Long), les cônes M ( aux moyennes Medium) et les cônes S aux courtes (Short)
Localisation des gènes des pigments chez l'homme
Les pigments photosensibles sont des protéines codées par des gènes. Le déséquilibre entre les sexes atteints de daltonisme s'explique par la présence des gènes codant pour les pigments sensibles au vert (moyennes longueurs d'ondes, pigment "vert", ou cônes M) et au rouge (grande longueur d 'onde, pigment "rouge" ou cônes L) sur le chromosome X. Si un homme XY porte un allèle déficient du gène du pigment M, ce dernier s'exprime et l'homme est dyschromate vert moins, alors qu'une femme devrait être homozygote pour cet allèle pour présenter le même phénotype.
Le fait que les gènes des pigments sensibles au vert et au rouge soient sur le chromosome X est connu depuis longtemps.
Les données nouvelles récemment acquises sur l'ensemble des gènes des pigments l'ont été grâce à la technique d'hybridation de l'ADN. A partir d'une sonde ADN du gène de la rhodopsine bovine (la rhodopsine est le pigment sensible à la luminance des bâtonnets) il a été trouvé différentes séquences fixant fortement -gène de la rhodpsine humaine-ou faiblement -gènes des pigments des cônes- cette sonde, selon leurs degrés de similitude dans leurs séquences nucléotidiques.
Les localisations suivantes ont été déterminées :
| PIGMENT | LOCALISATION DU GENE | Type de pathologie en cas d'absence du pigment | |
| Rhodopsine (bâtonnet) | chromosome 3 | ||
| Iodopsine "pigment rouge" (cônes L) | chromosome X | protanopie | |
| Iodopsine "pigment vert" (cônes M) | chromosome X | deutéranopie | |
| Iodopsine "pigment bleu" (cônes S) | chromosome 7 | tritanopie |
Si l'on admet que la similitude de ces séquences génétiques reflète une parenté ente ces gènes , la comparaison de ces dernières ou des séquences polypeptidiques des protéines correspondantes (40% de similitude entre séquences en acides aminés de la rhodopsine et des iodopsines) permet de se faire une idée de l'histoire évolutive de cette famille de gènes.
lien vers le logiciel phylogène
Les protéines des pigments rouge et vert se ressemblent beaucoup, elles ne diffèrent que par 15 acides aminés sur les 364 (les séquences nucléotidiques de leur gènes sont identiques à 98%), ce sont donc des gènes qui dérivent d'un gène ancestral récent. Les singes du Nouveau Monde n'ont sur leur chromosome X qu'un seul gène de pigment visuel (ils sont dichromates car possèdent deux types de cônes seulement : des cônes sensibles au bleu et des cônes sensibles à la zone rouge-vert), alors que les singes de l'Ancien Monde, plus proches de l'homme en possèdent deux. L'apparition du second gène porté par le chromosome X (par duplication et mutation du gène unique initialement présent sur le X) se serait ainsi produite après la séparation des continents africain et sud-américain, c'est à dire il y amoins de 40 millions d'années.
Remarque: les études expérimentales ont montré que 7 acides aminés jouent un rôle fondamental dans la sensibilité de la molécule d'iodopsine, en particulier 3 acides aminés dont les codons sont sur l'exon 5 du gène (acide aminés N° 277, 285 et 309). Si on remplace l'acide aminé ALA par THR au site 285, on décale de 14 nanomètres vers les grandes longueurs d'onde le pic de sensibilité du pigment (par exemple pour le pigment vert, le pic passe de 535 nm ou vert intense, à 549 nm ou vert-jaune).
Les multiples gènes visuels du X
On s'est récemment aperçu que les gènes des pigments visuels rouge et vert se succèdent sur le chromosome X (en accord avec leur formation par duplication d'un gène ancestral). Cet arrangement "en tandem" permet d'expliquer le fait étonnant que le nombre de gènes des pigments portés par le X diffère d'un individu à un autre.
Les échanges de fragments chromosomiques intra- et intergéniques par crossing-over (entre les deux X) et la multiplicité des gènes déjà notés par Jeremy Nathans (Université de Baltimore, USA) rendent compte du grand polymorphisme génétique de la vision colorée, qui comporte même à l'état normal, des gènes hybrides (entre ceux des pigments rouge et vert).
Dans l'espèce humaine, le nombre de gènes visuels portés par le X est encore discuté mais on admet
- qu'il n'y aurait qu'un seul gène du pigment rouge
- alors qu'on a décrit jusqu'à 9 gènes du pigment vert, avec 1 à 3 exemplaires de gène du pigment vert en moyenne sur le X, dont seul le premier (celui qui jouxte le gène du pigment rouge) est lu
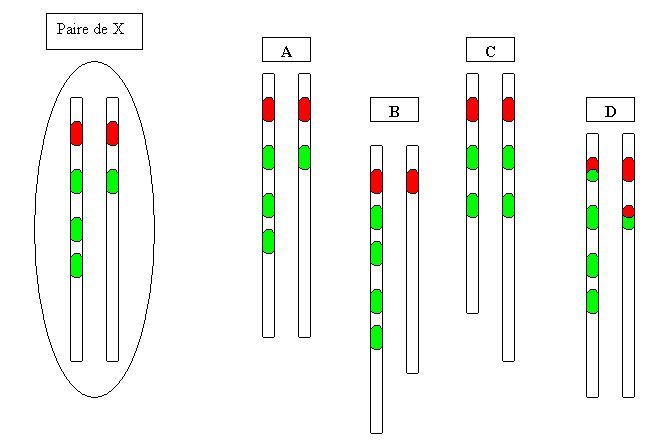 |
|
A partir des gènes des pigments rouge et vert tels que portés par la paire de chromosomes X schématisée à gauche, on peut obtenir les différentes paires représentées à droite, dont chaque chromosome passera dans un gamète et transmettra à la génération suivante diverses combinaisons de gènes |
Répartition statistique des déficits de la vision colorée chez l'homme (sexe masculin)
|
|
Classification |
Nom |
Etiologie |
Incidence en % chez les hommes (sexe masculin) |
|
DEFICITS
CONGENITAUX
|
Anomalie
Anomalie de la structure d’un des pigments en traînant une modification de sa sensibilité |
Protanomalie
|
pigment des cônes L anormal
|
1,3
|
|
Deuteranomalie
|
pigment des cônes M anormal
|
5,0
|
||
|
Tritanomalie
|
pigment des cônes S anormal
|
0,001
|
||
|
Dichromatopsie
Présence de deux types de cônes seulement
|
Protanopie
|
Cône L absent
|
1,3
|
|
|
Deuteranopie
|
Cône M absent
|
1,2
|
||
|
Tritanopie
|
Cône S absent
|
0,001
|
||
|
Monochromatopsie
Vision achromatique; pas de perception des couleurs
|
Typique
|
Tous les types de cônes absents
|
0,00001
|
|
|
Atypique ou incomplète
|
Deux types de cône absents
|
0,000001
|
||
|
DEFICITS |
Tritanopie
|
|
Maladie de la couche rétinienne externe
|
|
|
Deficit protan-deutan
|
|
Maladie de la couche rétinienne interne
|
|
|
|
VISION
NORMALE
|
|
|
Trois types de cônes présents
|
91,2
|
- Pour aller plus loin Détection des anomalies de la vision des couleurs (cas clinique)