Exploitation pédagogique
Dans le libellé du programme de TS, parmi les mécanismes de diversification du vivant à envisager , il est indiqué : hybridation suivie de polyploïdisation. Cela sous-entend que dans l’étude de la polyploïdie, on privilégie, l’allopolyploïdie. Les blés cultivés sont des allopolyploïdes et peuvent donc servir de support à cette étude. Pour l'aborder, on dispose :
- De figures de mitoses et de méioses (métaphases) de trois espèces de blé associées à un court texte de présentation ;
- Des caryotypes des 3 mêmes espèces de blé ;
- Quelques données sur la localisation chromosomique des gènes des gluténines et gliadines chez Triticum aestivum ;
- Schématisations de l’histoire évolutive des blés.
1 - Le nombre de chromosomes des espèces de blé
- La figure de métaphase d’une mitose chez Triticum monococcum montre distinctement 14 chromosomes ; celle de métaphase de méiose 1 montre 7 paires de chromosomes appariés. Cela conduit à la formule chromosomique 2n=14.
- La figure de métaphase de mitose de Triticum turgidum (durum) montre 28 chromosomes et celle de méiose 1 montre 14 paires de chromosomes appariés. On arrive dont à la formule chromosomique 2n=28.
Enfin, la photo de métaphase de mitose de Triticum aestivum permet plus difficilement de compter le nombre de chromosomes : 42 mais sur la photo de méiose 1 on distingue nettement 21 paires de chromosomes appariés. La formule chromosomique est donc 2n=42 chromosomes.
Avec ces documents, on ne peut conclure à la polyploïdie des blés dur et tendre car le comportement des chromosomes à la méiose chez ces deux espèces est typique de celui d’une espèce diploïde.
On peut seulement constater que les trois espèces de blé diffèrent par leur nombre de chromosomes et que ce nombre est un multiple de 7. Est-ce que cela est dû au hasard ou à un mécanisme à l’œuvre lors de la formation des deux espèces de blé dur et tendre ?
2 - Les caryotypes des trois espèces de blé
L’objectif est d’amener les élèves à s’interroger sur ces caryotypes à partir de leurs connaissances sur les caryotypes en général et des constats résultant de l’analyse des figures de mitose et méiose.
Le caryotype de Triticum monococcum présente toutes les caractéristiques d’un caryotype classique. Il s’agit donc du caryotype d’une espèce diploïde.
En revanche les caryotypes du blé dur et du blé tendre présentent une originalité. Celui du blé dur montre deux groupes de 7 chromosomes, désignés par les lettres A et B. Dans chaque groupe, chaque chromosome est représenté en deux exemplaires ce qui est habituel (chromosomes homologues). Le caryotype du blé tendre montre, outre les deux groupes de chromosomes A et B, un troisième groupe de 7 chromosomes, D.
Chez une espèce comme Triticum turgidum où le nombre de chromosomes est de 28, les chromosomes du caryotype devraient être numérotés de 1 à 14 et chez le blé tendre (42 chromosomes) de 1 à 21. Ce n’est pas le cas. On identifie seulement 7 chromosomes ayant chacun un représentant dans les groupes A et B (A1 et B1, A2 et B2, etc.) chez le blé dur, et un représentant dans les groupes A, B et D chez le blé tendre. Ainsi, on voit que le Triticum turgidum a 4 exemplaires de chaque chromosome et est donc tétraploïde ; Triticum aestivum a 6 exemplaires de chaque chromosome et est donc hexaploïde.
3 – La notion de chromosome homéologue
L’interprétation précédente suppose que les chromosomes 1 des trois groupes A, B et D sont homologues donc portent les mêmes gènes (et de même pour les chromosomes 2,3, et 4….7). Cette interprétation peut être testée à l'aide du document présentant la localisation chromosomique de quelques gènes. On constate que sur les chromosomes A1, B1 et D1 on trouve les gènes GLU et GLI1. Sur le chromosome 2 le gène Ppd1 et ainsi de suite. Ceci confirme qu'il s'agit de chromosomes homologues. Toutefois, et ce n'est pas uniquement dû au caractère partiel de la carte génétique utilisée, tous les gènes ne sont pas tripliqués. L’homologie des chromosomes A, B et D d'un groupe n’est donc pas identique à celle des chromosomes homologues au sens classique du terme. On dit qu’ils sont homéologues. L'homologie ne se traduit donc pas par une identité totale des gènes portés par les chromosomes.
La comparaison avec Anagène des séquences nucléiques des gènes GLU-A et GLU-B montre 72,3% d'identités. Celle des gènes GLU-A et GLU-D 77% d'identités, celle de GLU-B et GLU-D 73,3% d'identités. Ces pourcentages confirment que les gènes GLU-A, GLU-B et GLU-D sont homologues. Cependant, les différences dans les séquences des gènes sont nettement plus importantes que celles que l'on trouve habituellement entre les allèles d'un gène. Cela complète la notion de chromosomes homéologues.
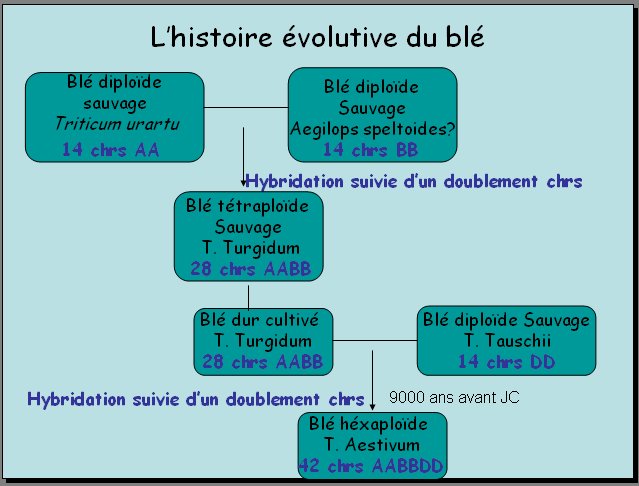
4 – Histoire évolutive des blés
Cette histoire doit rendre compte de la présence de 2 ensembles de chromosomes homéologues chez Triticum turgidum et de 3 ensembles chez Triticum aestivum.
Les blés sauvages, comme les graminées qui en sont proches, possèdent tous 14 chromosomes (2n=14), génome de base d'un blé. Par rapport à ces blés, les caryotypes de T. turgidum et de T. aestivum possèdent respectivement 1 et 2 génomes de base en plus. A partir de ce constat, on peut demander aux élèves d'émettre des hypothèses concernant l'origine de ces génomes supplémentaires. Les documents fournis permettent de tester ces hypothèses ou de donner lieu à une analyse directe pour en extraire une explication possible.
La première schématisation indique qu'une hybridation entre deux espèces de blés à 14 chromosomes (T. monococcum et A. speltoides) est à l'origine du T. turgidum ; une hybridation ultérieure entre T. turgidum et une autre espèce de blé à 14 chromosomes (T. tauschii) est à l'origine de T. aestivum. Le génome B provient donc de T. speltoides et le génome D de T. Tauschii.
La comparaison des séquences des gènes GLU de T. monococcum, T. speltoides et T. Tauchii confirme l'implication de ces espèces dans l'histoire évolutive des blés. En particulier, les séquences de GLU-D de T. aestivum et GLU de T. tauschii présentent près de 88% d'identités. C'est en accord avec l'implication relativement récente de T. tauschii (8000 à 9000 ans).
On comprend donc que les génomes A, B et D présentent une forte similitude car ils proviennent d'espèces proches mais qu'en même temps ils aient des différences (explication de l'homéologie).
En considérant la première hybridation, on peut engager les élèves à faire une critique de l'explication avancée. A partir de leurs connaissances sur la méïose, ils doivent dire que T. monococcum produit des gamètes à 7 chromosomes, ainsi que T. Speltoides. Les zygotes résultant de cette hybridation, et donc les plants de T. turgidum devraient avoir 14 chromosomes. En outre, puisque les chromosomes des deux espèces sont homéologues, ils devraient mal s'apparier et donner naissance à des gamètes anormaux dont l'hybride devrait être stérile. La seule hybridation est donc insuffisante pour expliquer le caryotype de T. turgidum.
La deuxième schématisation fournit une explication : hybridation suivie d'un doublement des chromosomes. Ce doublement peut avoir lieu au cours d'une mitose où il y a un comportement normal des chromosomes, mais où il n'y a pas de division du cytoplasme (ce qu'on peut provoquer artificiellement avec la colchicine). Cette mitose anormale peut se dérouler dès le développement de la cellule oeuf ou plus tardivement dans des organes qui contribueront à la formation des fleurs. Chez un tel hybride, la méiose se déroule normalement et conduit à des gamètes viables à 14 chromosomes. La même explication vaut pour la deuxième hybridation. Cette explication correspond à la phrase du programme : hybridations suivies de polyploïdisation.
Remarque : Une autre explication est théoriquement possible : les deux espèces qui s'hybrident produiraient des gamètes à 2n chromosomes (14) par suite de méiose anormale. L'hybride résultant des gamètes de ce type seraient directement tétraploïdes et fertiles. Il semble que ce mécanisme ne soit que rarement en jeu dans l'allopolyploïdie. En revanche, c'est le mécanisme le plus fréquent dans l'autopoplyploïdie.
5 - Bilan
On peut demander aux élèves de compléter les schémas en indiquant les génomes des différentes espèces de l'histoire. A, B et D représentent conventionnellement les génomes haploïdes de T. monococcum, T. speltoides et T. tauschii. Ils devraient aboutir au résultat suivant :