
La génétique de la tolérance au lactose
La génétique de la tolérance au lactose
(Dossier conçu dans le cadre du programme 2011. Révisé et adapté au nouveau programme de première spécialité SVT)
1 - Les phénotypes associés à la digestion du lactose
2 - Fréquence des phénotypes dans la population mondiale
3 - Déterminisme génétique de la différence phénotypique
4 - Un polymorphisme différent suivant les populations
5 - Le phénotype ancestral : LP ou LNP ?
6 - Datation de la mutation LP -13910T
7 - Les marques d’une sélection positive
8 - Une évolution biologique influencée par la culture
- Hypothèse
- Données sur les populations africaines et du Moyen-Orient
- Les débuts de l’élevage du bétail en Europe
- Les facteurs de sélection du phénotype LP
- Nouvelles hypothèses. Nature, juillet 2022.
1 - Les phénotypes associés à la digestion du lactose
Le lactose, principal glucide du lait, est un disaccharide formé par l’union d’une molécule de glucose et de galactose. Son absorption nécessite au préalable une hydrolyse réalisée par la lactase, enzyme de la bordure en brosse des cellules de l’épithélium intestinal.
Les adultes humains se répartissent en deux phénotypes en ce qui concerne l’aptitude à digérer le lactose. Les uns n’ont qu’une aptitude très faible à digérer le lactose car ils ne produisent plus de lactase (ou très peu). Ils sont dits « lactase non persistants » ou intolérants au lactose. Les autres dits « lactase persistants » gardent l’aptitude à digérer le lactose durant toute leur vie car leurs cellules intestinales continuent à produire de la lactase. Il faut bien voir que tous ces individus LP ou LNP durant les premières années de la vie produisaient de la lactase.
Chez les individus au phénotype « lactase non persistant », les manifestations d’intolérance au lactose débutent généralement vers 3-5 ans et se traduisent par un ballonnement abdominal, des douleurs abdominales, des borborygmes et, dans les cas les plus nets, des diarrhées.
2 - Fréquence des phénotypes dans la population mondiale
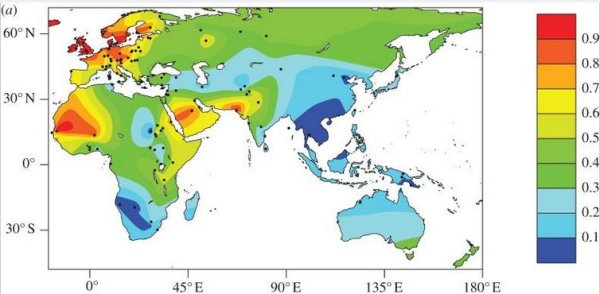
D'après : Evolution of lactase persistence: an example of human niche construction. Philos Trans R Soc Lond B Biol Sci. 2011 Mar 27; 366(1566): 863–877.
La fréquence du phénotype LP à l'échelle mondiale est estimée à 35% mais varie considérablement suivant les populations. Les plus fortes fréquences sur le continent européen sont observées dans le nord-ouest de l'Europe, en particulier dans les îles britanniques et la Scandinavie où elles varient entre 89% et 96%. On constate un déclin du phénotype LP du nord au sud et de l'ouest à l'est de l'Europe. Dans l'est de l'Asie, la fréquence est très faible. En Afrique, la distribution est hétérogène avec de forts contrastes entre populations voisines.
Le problème est d'essayer d'expliquer cette variabilité des phénotypes LP et LNP à l'échelle mondiale. Pour cela il faut faire le point sur le déterminisme génétique de ces phénotypes.
3 - Déterminisme génétique de la différence phénotypique
Les études familiales ont montré que les différences phénotypiques sont héréditaires et dues à un seul gène qui a été identifié comme celui qui code la lactase. Le gène de la lactase humaine, situé sur le chromosome 2, comporte 17 exons et code pour un polypeptide de 1927 acides aminés. Le phénotype lactase persistante est dominant.
Comparaison des séquences codantes du gène de la lactase
On peut comparer les séquences de la région strictement codante du gène de la lactase chez les membres d’une famille dont les deux parents ont le phénotype LP et qui ont deux enfants l’un LP, l’autre LNP.
Télécharger le fichier : Famille-LP-LNP.edi
La comparaison des séquences des allèles de ce gène chez les 4 membres de la famille ne montre aucune différence. Ce n’est donc pas une différence au niveau de la région codante du gène qui explique la différence phénotypique LP-LNP. On pouvait d’ailleurs le suspecter puisque les personnes LNP produisent une lactase fonctionnelle durant les premières années.
Régulation de l’expression du gène de la lactase
ll faut donc rechercher une autre explication au phénotype LNP. Puisque la lactase est produite pendant les premières années puis cesse de l’être, c’est donc qu’il y eu un changement dans l’expression du gène. A partir des connaissances sur les modalités de l’expression des gènes, on peut émettre diverses hypothèses :
- changement au niveau de la transcription ;
- changement au niveau de la traduction.
Pour tester la première hypothèse, les chercheurs, à partir de biopsies intestinales, ont recherché les ARN messagers du gène de la lactase chez des individus « lactase persistants » et « lactase non persistants ». Chez les premiers ils ont mis en évidence l’ARNm du gène de la lactase alors qu’il était absent chez les seconds. La différence phénotypique est donc due à une différence dans l’expression du gène et plus précisément dans sa transcription. Chez les « lactase non persistants », le gène s’exprime à la naissance mais cesse de s’exprimer à partir de 3-5 ans ; chez les « lactase persistants », il continue à s’exprimer durant toute la vie.
Pour aboutir à cette conclusion, on peut exploiter le fichier de séquences relatives à l’ARNm chez les personnes LP et LNP. Bien entendu, on ne peut traduire l’arrêt de production d’ARNm que par l’absence d’indication de séquence chez les personnes LNP après 5 ans.
Télécharger le fichier : ARNM-LP-LNP.edi
On introduit là l’idée qu’une différence phénotypique est liée à une différence dans la transcription du gène et non à une différence dans la séquence codante. C'est une notion du programme de terminale qui indique : « S’agissant des gènes impliqués dans le développement, des formes vivantes différentes peuvent résulter de variations dans la chronologie et l’intensité d’expression de gènes communs ». Certes, le gène de la lactase n’est pas un gène impliqué dans le développement mais il montre simplement qu’un changement dans la chronologie de l’expression d’un gène peut se traduire par une différence phénotypique.
Il s’agit de trouver la différence génétique (monogénique) qui est à l’origine du fait que chez certaines personnes, la transcription du gène de la lactase a lieu pendant toute la vie, alors que chez d’autres elle s’arrête vers 5 ans.
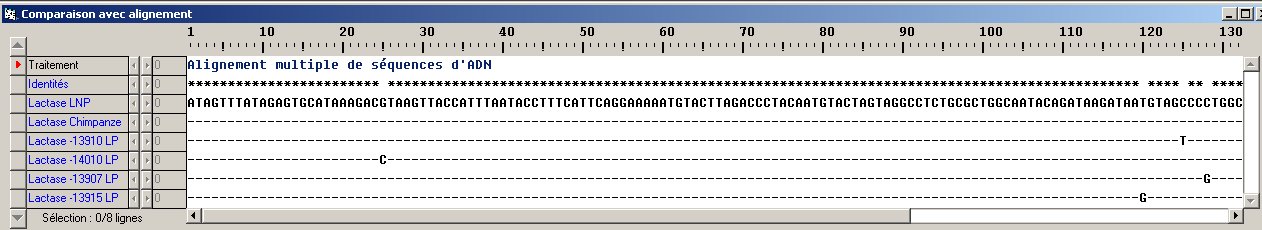
Cela implique d’enrichir la notion de gène en précisant qu’outre la séquence transcrite, il existe à proximité de celle-ci des séquences non transcrites mais qui affectent l’expression du gène. Dans le cas de la lactase cette zone régulatrice est située en amont du gène. On a mis en évidence que chez tous les individus européens de phénotype LNP, on trouve dans cette zone régulatrice, à la position -13910 (par rapport au 1er nucléotide transcrit), le nucléotide C (la paire C-G), alors que les personnes LP possèdent au même site le nucléotide T (la paire T-A).
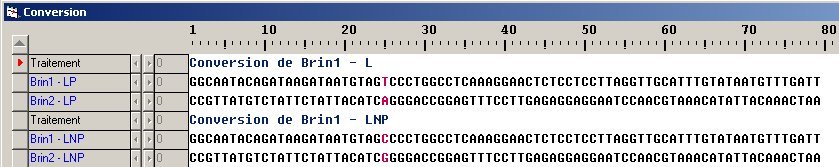
- Le fichier (Reg-Famille-LCT.edi) permet de comparer la séquence régulatrice chez les membres de la famille précédemment étudiée. La comparaison des séquences de cette région révèle donc l’association entre le génotype C/G en -13910 et le phénotype LNP.
Bien entendu, il est bon d’indiquer que les analyses génétiques réalisées sur d’autres familles européennes ont confirmé cette association.
Remarque : on se contente d'indiquer la différence génétique au niveau d'un seul brin et on parle donc de l'allèle -13910C et de l'allèle -13910T.
Des expériences de transgenèse ont confirmé l'implication du polymorphisme -13910 C/T dans la différence phénotypique. Cf. Informations scientifiques.
4 - Un polymorphisme différent suivant les populations
L’identification de l’allèle -13910T au début des années 2000 résulte de recherches effectuées sur les populations européennes. Les chercheurs ont ensuite analysé le génome des individus LP dans les populations africaines et du Moyen-Orient où ce phénotype est assez fréquent. Les deux cartes ci-dessous illustrent l’une la fréquence du phénotype LP et l’autre la fréquence de l’allèle -13910T dans l’ancien monde.
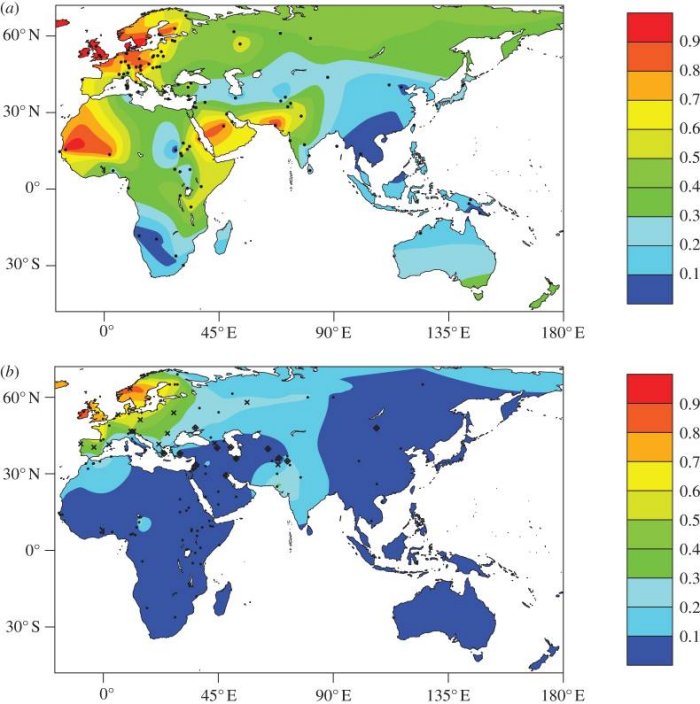
D'après : Gerbault et al. Evolution of lactase persistence: an example of human niche construction. Philos Trans R Soc Lond B Biol Sci. 2011 Mar 27; 366(1566): 863–877.
Si on constate en Europe une excellente corrélation entre la distribution du phénotype LP et celle de l’allèle -13910T, il n’en n’est pas de même en Afrique et au Moyen-Orient. Autrement dit, l’allèle -13910T ne semble pas l’agent causal du phénotype LP dans ces régions.
Une explication possible est que ce sont d’autres allèles de la séquence régulatrice du gène de la lactase qui sont à l’origine du phénotype LP chez les africains. En 2007, Tishkoff et al (Convergent adaptation of human lactase persistence in Africa and Europe. Nature Genetics 39, 31 - 40 (2006). Published online: 10 December 2006) rapportent avoir mis en évidence trois autres allèles dans les populations de l’Afrique de l’Est (Soudan, Kenya et Tanzanie) où le phénotype LP est fréquent. Ces allèles sont : -14010C (fréquent au Kenya et en Tanzanie chez les Massaï), -13915G (assez fréquent au Kenya) et -13907G (présent au Kenya et fréquent chez les Beja du nord du soudan). On constate que ces positions sont situées à proximité de -13910T trouvée chez les européens, dans un espace d’une centaine de bases. Cela permet de préciser ce qu’est une région régulatrice d’un gène dont des mutations ponctuelles différentes peuvent avoir le même impact sur l’expression du gène.
Le fichier de séquences (Allèles lactase.edi) comprenant les 4 allèles associés au phénotype LP permet, au terme d’une comparaison des séquences, d’établir les trois allèles africains et leurs polymorphismes -14010G/C ; -13915T/G et -13907C/G. Le premier nucléotide de chaque séquence est situé à la position -13934 par rapport au site d'initiation de la transcription du gène de la lactase.
La figure ci-dessous illustre la distribution du phénotype LP dans l’ancien monde prédite à partir des données sur la fréquence des allèles LP autres que -13910T. Par rapport à la seule prise en compte de la distribution du seul allèle -13910T, celle des 3 autres allèles rend mieux compte de la distribution du phénotype réellement observée en Afrique et au Moyen-Orient. Malgré tout, on constate des différences : ces 3 allèles ne permettent pas d’expliquer la fréquence du phénotype en Afrique de l’Ouest. Probablement qu’un ou plusieurs autres allèles sont à découvrir.
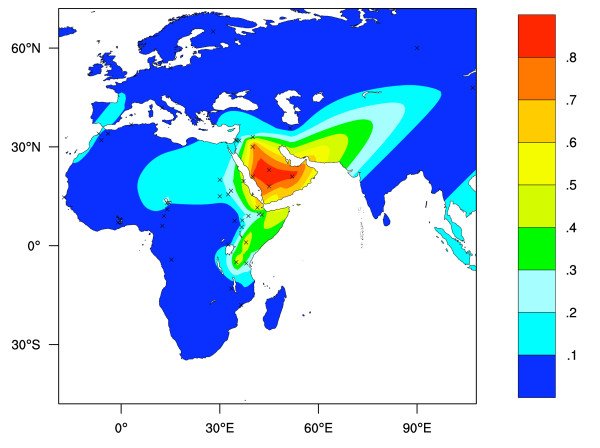
D'après : Yuval Itan et al. : A worldwide correlation of lactase persistence phenotype and genotypes. BMC Evol Biol. 2010; 10: 36. Published online 2010 Feb 9.
5 - Le phénotype ancestral : LP ou LNP ?
65% environ de la population mondiale est de phénotype LNP mais cela ne prouve pas que c’est le phénotype ancestral et LP le phénotype dérivé. Les données suivantes permettent d’argumenter.
- La quasi-totalité des mammifères adultes ne consomment pas de lait et surtout ne synthétisent plus de lactase.
- On a séquencé la région régulatrice du gène de la lactase chez le Chimpanzé, donc une séquence homologue de la séquence humaine. La comparaison de cette séquence avec les 4 allèles LP humains (Fichier Allèles LCT Humains et Pan.edi) montre qu'aux 4 polymorphismes humains on trouve chez le chimpanzé le nucléotide présent chez les individus LNP : -14010G ; -13915T ; -13910C ; -13907C. Donc la séquence LNP était présente chez l'ancêtre commun à l'Homme et au Chimpanzé et les 4 allèles LP résultent chacun d'une mutation de la séquence régulatrice dans les populations humaines.
6 - Datation de la mutation LP -13910T
Dans une publication de 2007, Burger et al. relatent les résultats obtenus à partir de l’ADN extrait de 8 squelettes de sites archéologiques d’Allemagne, de Hongrie, de Pologne et de Lituanie. L’âge de ces os est compris entre 5800 et 5000 ans avant J.C. Ils ont recherché la présence de l’allèle -13910T dans chacun des échantillons. A l’aide du fichier (Table 1. Samples, culture, archaeological or radiocarbon dating, lactase genotype, and mtDNA haplotype), on peut découvrir que les 8 individus étaient homozygotes au SNP -13910 (SNP : Single Nucleotide Polymorphism) et possédaient la paire de nucléotides C/C.
|
Culture |
Archaeological or radiocarbon dating |
Génotype-13910 |
|
Neolithic Linear Pottery |
5500–5000 B.C. |
C//C |
|
Neolithic Körös |
5840–5630 B.C. |
C//C |
|
Middle Neolithic Narva |
5580 ± 65 B.C. |
C//C |
D'après : J. Burger et al. Absence of the lactase-persistence-associated allele in early Neolithic Europeans. PNAS vol. 104 no. 10 > J. Burger, 3736–3741
- Otzi est une momie qui a été découverte en 1991 dans le massif de l’OtzaÏ, à la frontière Italo-Autrichienne, par un couple de randonneurs allemands. Il gisait à 3210 m d’altitude. On a pu dater sa mort : 5300 ans.
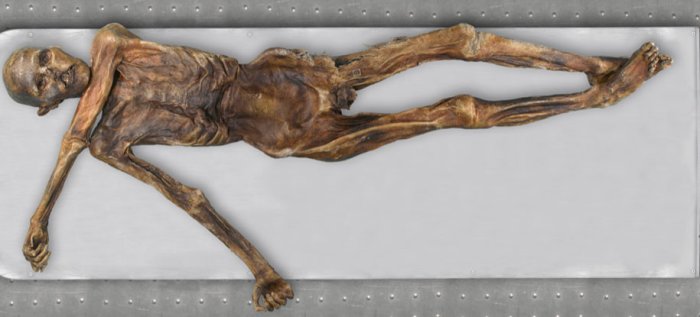
Le squelette d'Otzi (Homme des glaces).
D'après : New insights into the Tyrolean Iceman's origin and phenotype as inferred by whole-genome sequencing. Andreas Keller et al. Nature Communications 28 February 2012.
L’analyse de son ADN a révélé qu’au site -13910 de la région régulatrice du gène de la lactase, il possédait le génotype C//C.
Bilan
L’exploitation de ces différents types de données débouche sur l’idée que le phénotype LNP est ancestral et le phénotype LP dérivé. Puisque les tests de tolérance/intolérance au lactose et les biopsies intestinales (recheche de la lactase) montrent que les mammifères adultes et notamment les grands Singes ont tous le phénotype LNP, comme 65% des humains, cela signifie que l’ancêtre commun à l’Homme et au Chimpanzé possédait aussi ce phénotype et que le phénotype LP est apparu dans la lignée humaine. Cette conclusion est confortée par les données génétiques car, à tous les sites de la région régulatrice du gène de la lactase impliqués dans la réalisation du phénotype LP, les humains LNP possèdent les mêmes nucléotides que le Chimpanzé. Les données sur l’ADN ancien des européens du néolithique semblent indiquer qu’il y a 5000 à 6000 ans les européens d’Europe centrale avaient le phénotype LNP alors qu’actuellement la majorité ont le phénotype LP. Certes, l’analyse n’a porté que sur 8 individus du Néolitique et c’est peu pour déterminer les caractéristiques d’une population, néanmoins cela incite à penser que le phénotype LP était peu répandu à cette époque en Europe. Cela est confirmé d’ailleurs par la momie Otzi et par les conclusions d’une publication de 2014 sur l’analyse de l’ADN ancien extrait de sites archéologiques de Hongrie (Genome flux and stasis in a five millenium transect of European prehistory; Cristina Gamba et al. Nature October 2014).
En conclusion, on peut dire que le phénotype LP, phénotype dérivé, résulte de mutations survenues dans la région régulatrice du gène de la lactase dans diverses populations. Ces mutations indépendantes les unes des autres, ont eu pour effet de prolonger chez l’adulte l’expression du gène de la lactase et donc de conduire au même phénotype. C’est un exemple d’évolution convergente dans l’espèce humaine. Il reste à comprendre les facteurs qui ont contribué à l’expansion de ces mutations dans les populations.
7 - Les marques d’une sélection positive
A partir de techniques complexes, les chercheurs ont fait des estimations sur les dates d’apparition des diverses mutations à l’origine du phénotype LP. Ainsi, pour la mutation -13910T des européens, elle serait apparue entre -10000 ans et -5000 ans avant J.C. La datation pour les mutations africaines serait du même ordre, peut-être un peu moins anciennes. De toute façon, à l’échelle de temps de l’évolution, ce sont des mutations récentes. La seule dérive génétique ne permet pas d’expliquer qu’elles aient pu se répandre à un tel degré dans les populations en si peu de temps, notamment pour la mutation européenne qui est presque fixée dans les Iles britanniques et la Scandinavie. Cela suggère l’intervention de la sélection naturelle.
Il reste à envisager les facteurs ayant contribué à cette sélection positive des allèles LP et par là à l’évolution phénotypique des populations.
8 - Une évolution biologique influencée par la culture
Hypothèse
Le lactose est présent uniquement dans le lait. La possession d’allèles LP ayant pour unique effet de permettre de digérer le lactose à l'état adulte ne peut entraîner une sélection positive de ces allèles que si les populations en cause consomment du lait, donc pratiquent un élevage laitier. De là découle l’hypothèse que c’est la domestication du bétail, chèvres, brebis et vaches, etc., donc le passage d’une société de cueilleurs-chasseurs à une société de fermiers éleveurs qui a créé le nouvel environnement ayant permis l’évolution biologique du phénotype LNP au phénotype LP. L’évolution culturelle transmise de génération en génération a entraîné l’évolution biologique et sans doute que celle-ci en retour a influencé l’évolution culturelle.
Données sur les populations africaines et du Moyen-Orient
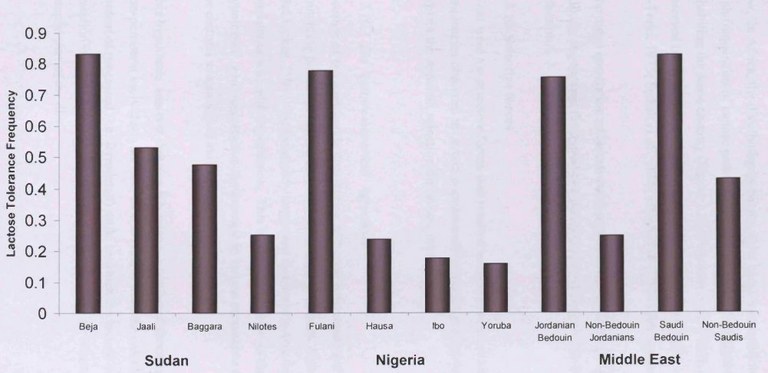
Les histogrammes ci-dessous renseignent sur la fréquence du phénotype LP dans les populations de deux pays africains et dans celles du Moyen-Orient.
D'après : Lactose digestion and the evolutionary genetics of lactase persistence. Catherine J. E. Ingram et All. Hum Genet (2009) 124:579–591
Une étude réalisée en Jordanie a révélé que dans la population de bédouins du désert au mode de vie nomade et qui consomment le lait de leur bétail (ovins, caprins et chameaux) la fréquence du phénotype LP était de 76%, alors que dans la population d’arabes non bédouins des zones urbaines et agricoles de Jordanie, la fréquence de LP n’était que de 25%. La même observation a été réalisée en Arabie saoudite. La population Beja du nord du Soudan à mode de vie nomade et pastoral et qui consomme le lait de son bétail est à 87% de phénotype LP alors que chez les Nilotes du sud du soudan dont l’alimentation repose beaucoup moins sur le lait et produits laitiers, la fréquence des LP est de 25,5%.
Chez les Masaï du Kenya et de Tanzanie (Afrique de l’est) qui mènent une vie semi nomade et qui élèvent du bétail et consomment en particulier du lait, la fréquence du phénotype LP est respectivement de 71% et 59% ; chez les Sandawe, population de chasseurs cueilleurs de Tanzanie, le fréquence du phénotype LP est 22%.
Ce qui ressort de manière systématique c'est la fréquence élevée des LP dans les populations qui consomment du lait.
Les débuts de l’élevage du bétail en Europe
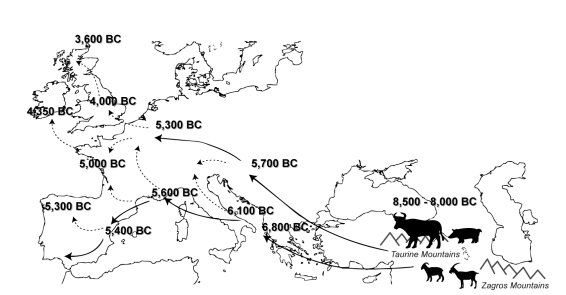
- Les informations fournies par les sites archéologiques indiquent que c’est au Proche-Orient qu’est apparue la domestication des animaux un peu avant -10000 ans. C’est le début de la révolution néolithique où certains groupes humains passent du statut de chasseurs-cueilleurs à celui d’agriculteurs-éleveurs. Les espèces animales domestiquées sont : la chèvre, le mouton, le porc et le bœuf ; elles le sont à partir des ancêtres sauvages vivant dans ces régions du Proche-Orient : l’aurochs (ancêtre du bœuf), la chèvre aegare, le mouflon oriental (ancêtre du mouton) et le sanglier. C’est aussi les débuts d’un mode de vie sédentaire avec création des premiers villages.
- Durant les millénaires qui suivent, la pratique de l’élevage se propage vers l’ouest comme l’illustre la carte ci-dessous. On voit que la diffusion de l’élevage en Europe s’est faite suivant deux grandes voies : une route méditerranéenne et une route plus septentrionale qualifiée de danubienne, qui passe par les Balkans et l’Europe centrale. Vers -4000 ans avant J.C l’élevage du bétail s’était répandu dans toute l’Europe, donc en 5 millénaires.
D'après : Leche the book. Page 39.
- Deux modèles sont proposés pour rendre compte de cette diffusion. Suivant le premier, en liaison avec un accroissement démographique, il s’agit de migration progressive d’éleveurs avec leur bétail. Cela est confirmé par les études génétiques indiquant que les bovins du néolithique étaient génétiquement très différents des aurochs européens et semblables aux aurochs proche-orientaux, ce qui plaide fortement pour un transport d’animaux. Suivant le second, il s’agit d’un processus de transmission de pratiques culturelles, les populations de chasseurs-cueilleurs d’Europe adoptant progressivement les modes de vie des éleveurs. Il est fort probable que les deux processus ont contribué à ce que finalement toute l’Europe soit peuplée uniquement par des éleveurs.
- Pendant longtemps, on a estimé que l’élevage pour produire du lait et le consommer était apparu plusieurs millénaires après le début de la domestication. Les travaux récents, notamment ceux des chercheurs du projet européen Leche, ont remis en cause cette conception. L’analyse des résidus organiques contenus dans les plus anciennes poteries connues en Turquie nord-occidentale a révélé la présence de lipides laitiers, signe que dès -9000 ans avant J.C l’élevage bovin laitier tenait une place importante dans cette région. Dans des sites archéologiques de Pologne, on a trouvé des poteries percées de trous datant de -7500 ans avant J.C interprétées comme des faisselles ayant servi à égoutter les caillots de lait pour faire du fromage. L’analyse des résidus organiques contenus dans ces poteries a révélé la présence de résidus adipeux et l’analyse chimique précisé qu’il s’agissait de lipides laitiers. Cela fait de ces reliques polonaises la plus ancienne trace connue de la fabrication du fromage dans le monde. Cela traduit que l’élevage à des fins de production de lait et de consommation de produits laitiers, a débuté rapidement après les débuts de la domestication.
Les facteurs de sélection du phénotype LP
- Le fait qu’en Afrique et au Moyen-Orient le phénotype LP soit plus fréquent dans les populations nomades élevant le bétail que chez celles dont l’alimentation dépend surtout de la cueillette et de la chasse corrobore l’hypothèse suivant laquelle l’élevage du bétail a créé un environnement entraînant un avantage sélectif aux allèles du gène de la lactase, maintenant l’expression du gène chez l’adulte. Chez les européens, l’expansion de l’allèle LP -13910 a eu lieu en même temps que l’élevage du bétail se généralisait, notamment celle de la production de lait, ce qui confirme aussi l’hypothèse. Certes, les populations européennes du néolithique d’il y a -5000 à -6000 ans, étaient sans doute majoritairement de phénotype LNP, bien que la production laitière ait débuté depuis 2 à 3 millénaires. Le fromage est nettement moins riche que le lait en lactose ce qui fait qu’il peut être consommé par les personnes LNP sans inconvénient majeur. C’est sans doute après que la consommation du lait par les adultes s'est répandue que l’avantage sélectif dû à sa consommation a entraîné l’expansion de l’allèle LP -13910. Il reste à préciser en quoi consiste cet avantage sélectif.
- Le premier avantage peut résider dans l’apport énergétique du lait. On estime que la production de lait par une vache préhistorique devait être entre 400 et 600 kg suite à une gestation. Après avoir soustrait la quantité de lait nécessaire au jeune veau, il reste 150 à 200 kg. Ceci est presque équivalent à l’apport énergétique obtenu à partir de la viande d’une vache. Cet apport énergétique qui nécessite de digérer le lactose pouvait être particulièrement important dans les périodes de disette entre les périodes de récolte des cultures céréalières et donc favoriser la survie des personnes LP.
- Un deuxième avantage sélectif fourni par le lait est en rapport avec l’assimilation du calcium. La vitamine D favorise l'absorption intestinale du calcium. Dans les régions nordiques où le rayonnement UV est faible pendant plusieurs mois de l’année (voir dossier pigmentation de la peau) la production cutanée de précurseurs de la vitamine D, sous l’action des UV, est réduite. Si l’apport alimentaire des éleveurs du néolithique fournissait peu de vitamine D, il en résultait des risques de rachitisme. Le lait en apportant le calcium et un peu de vitamine D pouvait contribuer à l’éviter. L’avantage sélectif fourni par la consommation du lait selon cette hypothèse n’est pas en relation directe avec la capacité à digérer du lactose, puisque ce sont d’autres constituants du lait qui sont en jeu. Il est toutefois en relation indirecte puisque les personnes LNP ne pouvant consommer du lait à cause de leur incapacité de digérer le lactose, ne bénéficiaient pas de l’apport vitaminique et en calcium du lait.
- Une autre explication est en rapport avec le climat. Dans les régions où sévit la sécheresse, le lait représente une source d’eau non polluée. Les personnes LNP ne pouvaient bénéficier de cette source d’eau et de plus, les diarrhées en cas de consommation de lait pouvaient entraîner une déshydratation pouvant être mortelle.
- Bien entendu les explications peuvent être différentes suivant les régions. En Afrique où le rayonnement UV est fort, il est peu probable que l’explication faisant intervenir la vitamine D ait joué un rôle important. Inversement, l’explication relative à l’apport d’eau a certainement eu peu d’impact dans les régions nordiques.
- Le phénotype LNP est ancestral. Le phénotype LP résulte de mutations intervenues entre 5000 ans et 10000 ans avant J.C. Les populations chez lesquelles le phénotype LP est devenu fréquent ont toutes une histoire de pratique de l’élevage. Le génome des individus LP montre des signatures d’une sélection positive. Tout cela fait qu’il y a un accord pour dire qu’au cours de l’histoire des populations, l’extension du phénotype LP résulte d’avantages sélectifs que fournissait la consommation du lait à condition de digérer le lactose. C’est un exemple où une innovation culturelle, l’élevage, en créant un nouvel environnement, l’apport de lait, a créé un avantage sélectif qui a entraîné une évolution phénotypique des populations. Probablement, la généralisation de l’aptitude à digérer le lait et donc à l’exploiter a eu des conséquences sur l’évolution des techniques d’élevage donc sur les pratiques culturelles.
- L’Homme par sélection a fait évoluer le phénotype des animaux laitiers. Inversement le lait produit par les vaches ou les chèvres a fait évoluer le phénotype LNP dans les populations humaines. C’est un exemple de coévolution dû à une sélection artificielle pour le bétail, et à la sélection naturelle pour les humains.
- Cela étant, il reste des points non résolus. On pourrait s’attendre à ce que la fréquence du phénotype LP soit la plus élevée dans les descendants des populations qui ont pratiqué en premier l’élevage, ce qui n’est pas le cas. En outre, il faudrait expliquer la diminution en Europe de la fréquence du phénotype LP en relation avec la latitude, alors que les populations d’Europe du sud ont aussi une tradition historique d’élevage. Les travaux des chercheurs du projet Leche ont entrouvert des pistes d’explication en mettant en évidence une régionalisation des pratiques laitières, de la consommation de lait ou de fromages, ou de laits fermentés (seule la consommation de lait peut fournir un avantage sélectif au phénotype LP). La régionalisation des pratiques est probablement à l’origine des pressions de sélection qui ont permis en Europe à la mutation -13910 de se généraliser très rapidement dans certaines régions et pas dans d’autres. Il suffisait par exemple qu’on ait plutôt consommé du lait liquide ici, du yaourt ou du fromage là, pour que les deux populations humaines aient suivi des évolutions génétiques différentes du point de vue du gène de la lactase.
Nouvelles hypothèses émises en juillet 2022
- Dairying, diseases and the evolution of lactase persistence in Europe. Richard P. Evershed et al. Nature. Published: 27 July 2022.
- Article en Français : Bien digérer le lait, un avantage évolutif en temps de crise.
" L’étude publiée dans Nature renverse certaines croyances sur la raison pour laquelle les humains ont développé la capacité de digérer le lactose à l’âge adulte. Plutôt que l’habitus alimentaire lié à la domestication, ce sont au contraire des facteurs de mortalité, tels que la famine et l’exposition aux maladies, qui expliquent le mieux l’évolution de notre tolérance au lactose en Europe."