
Arbres phylogénétiques et compréhension de l’évolution
Arbres phylogénétique et compréhension de l'évolution
Jean-Claude Hervé, septembre 2010
I – Le « Tree-thinking » n’est pas si évident…
La capacité à tirer des informations pertinentes sur les relations de parenté entre les espèces à partir d’un arbre phylogénétique est un objectif du programme de terminale S du début des années 2000. Certes, le terme de phylogénie apparaît dans les deux programmes précédents mais sans référence précise à la méthodologie cladiste. De même, la consultation d’ouvrages scolaires anglo-saxons, montre que l’analyse d’arbres phylogénétiques fait aujourd’hui partie intégrante des programmes. En outre la littérature scientifique est désormais riche en arbres phylogénétiques, et cela dans des domaines qui dépassent le cadre de la systématique (Exemple dans un domaine des sciences humaines). Le « tree thinking », pour reprendre l’expression des anglo-saxons, fait partie de la culture biologique de base, et amène à envisager les organismes dans une perspective historique.
Après avoir, pendant longtemps, mis l’accent sur les mécanismes évolutifs, les « process », l’enseignement de l’évolution accorde actuellement plus d’importance aux « patterns », à la reconstitution de l’histoire de la vie suivant une démarche scientifique rigoureuse. Les arbres phylogénétiques apparaissent ainsi comme la traduction visuelle du concept central de l’évolution, la "descendance avec modifications"de Darwin. C’est aussi une façon de structurer la diversité biologique.
Durant ces dernières années, plusieurs scientifiques et enseignants, principalement anglo-saxons, ont toutefois mis l’accent sur la fréquence des interprétations erronées d’arbres phylogénétiques, de la part du grand public, des lycéens, même après avoir eu un enseignement de biologie de l’évolution, et aussi parfois de scientifiques non spécialistes de la systématique. Ces fausses interprétations sont généralement dues à une conception spontanée et erronée de l’évolution, qui sert de guide à la lecture des arbres. Inversement, l’exploitation de certains arbres comme celui représenté ci-dessous, renforce si elle reste trop superficielle, les conceptions initiales des lycéens sur l’évolution.
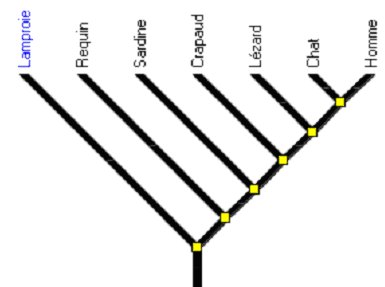
Figure 1. Exemple d'un arbre construit pour illustrer les difficultés de lecture
Ces arbres sont « déséquilibrés » par suite d’un biais dans la sélection des taxons et des caractères, ce qui conduit à indiquer des innovations évolutives uniquement sur la branche de droite à partir du nœud basal. Cela ne signifie pas qu’il faut s’interdire d’envisager de tels arbres avec les élèves car c’est sans doute un excellent moyen de connaître leurs représentations initiales, relatives à la lecture des arbres phylogénétiques.
II – Les erreurs liées à l’idée de plus ou moins grand degré d'évolution des taxons
A – La prévalence de « l’échelle des êtres »
Pendant des siècles, la notion d’échelle des êtres a joué un rôle central dans la pensée occidentale. Cette conception indique que l’univers est ordonné suivant une séquence linéaire commençant au monde inanimé des roches, les plantes viennent après, puis les animaux, les hommes, les anges, et finalement Dieu. Pour se limiter au monde vivant, l’échelle des êtres conduit à classer les êtres vivants suivant leur degré de perfection ; chaque niveau de l’échelle possède les propriétés des niveaux en dessous plus celles qui lui sont propres.
B – L'idée de « progrès évolutif »
Le concept d'échelle des êtres, prégnant dans un mode fixiste a été par la suite et jusqu’à très récemment introduit implicitement dans la perspective évolutive du monde vivant (voir l'échelle des êtres de Haeckel) alors que le seul arbre dessiné par Darwin est plus neutre. Il n’y a qu’à songer au résumé qu’on fait de l’histoire de la vie. Au départ apparaissent les organismes cellulaires dépourvus de véritable noyau, puis les eucaryotes unicellulaires, puis les organismes pluricellulaires, etc.
L’évolution se caractérise par l’apparition d’organismes de plus en plus complexes au fur et à mesure de son déroulement, le maximum de complexité se réalisant avec l’apparition de l’espèce humaine. C’est l’idée de progrès évolutif dénoncé par Stephen Jay Gould. C’est ce qui conduit à parler pour des espèces vivant actuellement d’organismes primitifs ou inférieurs, d’organismes évolués ou supérieurs.
En ce qui concerne le monde animal, la référence pour dire qu’une espèce est évoluée est l’espèce humaine : celles qui possèdent beaucoup de caractéristiques de l’espèce humaine sont "plus évoluées" que celles qui ne les ont pas. La très grande majorité des ouvrages universitaires de zoologie ou de botanique du 20ième siècle font usage de telles expressions (Par exemple, la 5ème édition d’un abrégé de biologie, parue en 1996 avait pour titre : Plantes supérieures : appareil végétatif).
Cette idée d’organismes plus ou moins évolués est profondément ancrée dans notre culture et sert spontanément de guide dans l’interprétation des arbres phylogénétiques. Il n’est d’ailleurs pas sûr que les termes que nous utilisons aujourd’hui dans notre enseignement ne renforcent pas, si on n’y prend pas garde, cette représentation du monde vivant. Pour éviter un jargon rebutant (apomorphe, plésiomorphe), on préfère utiliser les expressions : état dérivé , état ancestral d’un caractère. Mais les lycéens, si on n’attire pas leur attention, passent facilement de taxon possédant l’état primitif ( ou ancestral ) à taxon primitif, inférieur, voire à ancêtre. L’introduction récente dans Phylogène de la notion d’extra groupe (taxon utilisé par les scientifiques pour polariser les caractères), peut également conduire à cette confusion… si on n’y veille pas. Comment ne pas penser, au premier abord, que le taxon extra groupe qui possède tous les états ancestraux des caractères qui servent à établir des relations de parenté dans le groupe d’étude n’est pas primitif !
L’arbre sous forme de barreaux d’échelle montré précédemment (Figure 1) se caractérise par l’absence de branchements sur une des lignes issues du nœud basal et sur les ramifications latérales de la ligne principale. Les enquêtes menées par les chercheurs américains indiquent que de nombreux étudiants pensent qu’il n’y a pas eu d’évolution dans ces lignées. Cela les conduit à penser que le taxon à l’extrémité de la branche liée au nœud basal est l’ancêtre de tous les autres taxons et est donc peu évolué, primitif.
D’une façon plus générale, en rapport avec l’idée d’espèces plus ou moins évoluées, c’est l&rsqrsquo;idée qu’une espèce actuelle peut être l’ancêtre d’une autre espèce actuelle qui est sous jacente 1.
Cela fait songer à un essai 2 où S. J. Gould dit :
« Dans toute la biologie de l’évolution, je ne pense pas qu’il y ait d’erreur plus fondamentalement instructive ou plus fréquemment répétée qu’un certain raisonnement erroné au sujet des grands singes et de l’homme. Si l’évolution est vraie et si nous sommes effectivement issus des grands singes, alors pourquoi ces derniers sont-ils encore vivants ? ».
D’une façon beaucoup moins caricaturale et en considérant des groupes plus vastes, c’est un peu le même raisonnement qui faisait dire, jusqu’il y a peu, que les Batraciens sont issus des Poissons, les Reptiles des Batraciens, les Mammifères des Reptiles, etc. (voir aussi classification linnéenne et classification phylogénétique)
Un des objectifs de l’étude d’arbres phylogénétiques est de faire saisir qu’il n’y a pas de hiérarchie objective dans les taxons existant actuellement, que toutes les espèces sont évoluées, et présentent chacune une mosaïque d'états dérivés et d'états ancestraux de caractères, mosaïque variable d’un taxon à l’autre. Par exemple, l’ornithorynque était classiquement présenté comme un Mammifère primitif du fait des caractéristiques de sa reproduction. Mais il possède des états dérivés comme un bec corné ou un éperon venimeux sur les pattes arrières que ne possèdent pas les autres Mammifères. On peut certes attribuer une plus grande valeur à certains états dérivés (comme la possession d’un gros cerveau !) mais c’est là un jugement de valeur qui n’est pas du domaine des sciences.
C – Des activités pour combatte ce type d'erreurs
1 - Dans un premier temps destiné à explorer les conceptions initiales des élèves, il est possible de poser des « questions provocantes» du type : « Quelle est l’espèce la plus vieille ? La plus jeune ? la plus primitive ? la plus évoluée ? la plus complexe ? » à partir d'un arbre valide mais cristallisant des obstacles (figure 2) Bien entendu cet arbre ne fournit aucune information permettant de répondre à ces questions. Mais l’analyse des réponses des élèves à ces questions peut fournir des informations intéressantes sur leurs conceptions sur l’évolution et orienter les démarches à suivre par la suite. (Posée à nouveau à la fin d'un cours, ces mêmes questions devraient aboutir à des réponses du type " cette question n'a pas de sens...")
Figure 2. Un arbre caricatural (mais valide) pour faire ressortir les représentations initiales.
2 - Envisager d'emblée un exemple d’arbre où l’Homme sert d'extragroupe et se trouve situé à gauche sur l’arbre permet d'éviter les interprétations erronées. Par exemple, la sous-collection Diapsides permet d’établir des relations de parenté au sein de ce groupe grâce à des innovations que l'Homme ne possède pas (figure 3). La collection "Périssodactyles" de Phylogène peut aussi servir de support. à ce type d’activité.
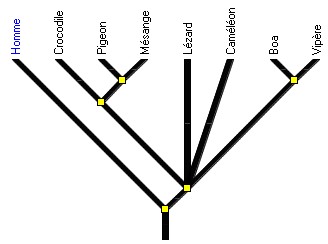
Figure 3. L'arbre des Diapsides. L'Homme (extragroupe) sert à définir les états ancestraux des caractères.
3 - Il paraît aussi nécessaire de proposer à l’analyse un arbre où il y a des ramifications dans toutes les branches de l’arbre (ou au moins un arbre où tous les taxons terminaux possèdent des états dérivés qui leur sont propres). Cet arbre illustrera ce qu’il est classique d’appeler évolution buissonnante, représentation qui traduit bien qu’il y a eu évolution dans toutes les lignées. Vouloir présenter uniquement des arbres « simplistes » avec peu de taxons et peu de caractères peut conduire à renforcer des idées erronées sur l’évolution.
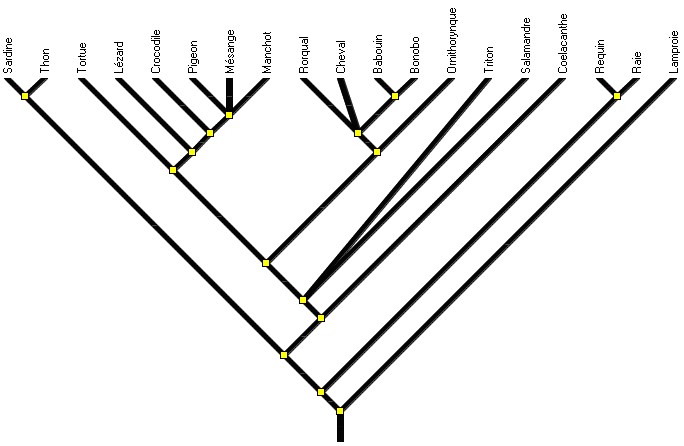
Figure 4 . L'arbre des Vertébrés montrant une évolution buissonnante.
Remarque : Le film "Espèce d'espèces" diffusé depuis l'an dernier et la sphère de parenté qu'il présente permet sans doute une vue plus parlante de l'évolution buissonnante.
Dans Phylogène, on trouve des collections dont l’étude n’est pas spécifiquement prévue au programme ; Cétacodontes, Téthythériens, Equidés. Des arbres élaborés à partir de ces collections peuvent être utilisés pour traduire l’existence d’une évolution dans toutes les lignées.
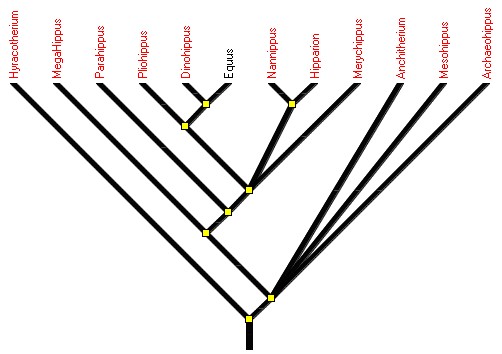
Figure 5. Evolution au sein du groupe des Équidés
4 - Une des améliorations récente de Phylogène est la possibilité de représenter les groupes monophylétiques sur les arbres construits sous forme de boîtes et de nommer les groupes emboîtés qui en résultent (figure 6).
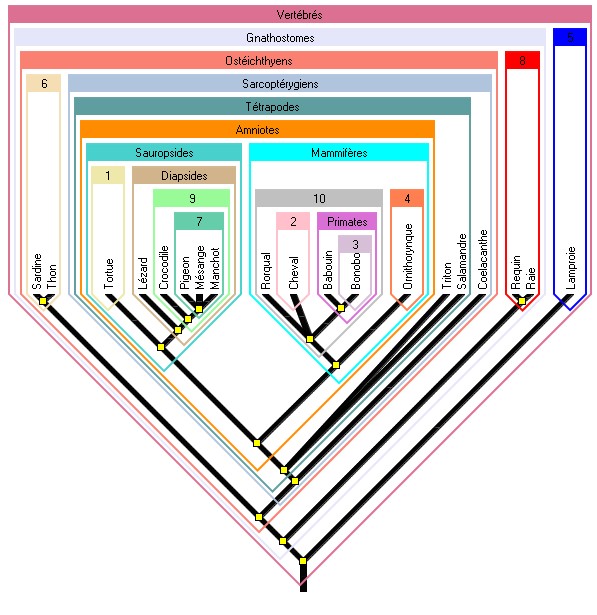
Figure 6. Arbre des Vertébrés dont les groupes monophylétiques sont nommés
Cela est en accord avec la phrase du programme de TS qui dit : « L’Homme est un Eucaryote, un Vertébré, un Tétrapode, un Amniote, un Mammifère, un Primate, un Hominoïde, un Hominidé, un Homininé ». C’est une profonde rupture avec la systématique ancienne qui faisait dériver des groupes évolués de groupes primitifs comme rappelé précédemment. Cela à condition qu’on utilise le langage adéquat : les Oiseaux ne sont pas issus des Dinosaures mais sont des Dinosaures ; les hommes ne sont pas issus des singes (compris comme Simiiformes) mais ce sont des singes.
La révolution dans la façon de penser l’évolution ne peut devenir effective que si on fait ce travail de classification emboîtée (aboutissant à "l’adresse de clades" - Clad address) pour plusieurs espèces. Phylogène le permet, notamment si on utilise la collection des Amniotes, ce qui peut donner lieu à des exercices adaptés.
5 - Ce travail sur les groupes emboîtés,en prolongement de l’étude des arbres, permet de situer chronologiquement l’ordre d’apparition des différentes innovations acquises par un organisme. Grâce à la fonction « Dater », si des taxons fossiles ont été choisis, on peut même proposer un âge absolu approximatif. C’est la structuration de la biodiversité dans une perspective évolutive.
Il peut être intéressant de faire remarquer que toutes les lignées d’un arbre menant aux espèces actuelles ont évolué, depuis la racine de l'arbre jusqu'à aujourd'hui, pendant le même temps. Même si les Téléostéens sont apparus bien avant les Primates, la durée d’évolution dans les lignées qui conduisent aux Téléostéens actuels et aux Primates actuels est la même (figure 7). Cette réflexion sur le temps d’évolution peut renforcer l’idée d’évolution dans toutes les lignées.
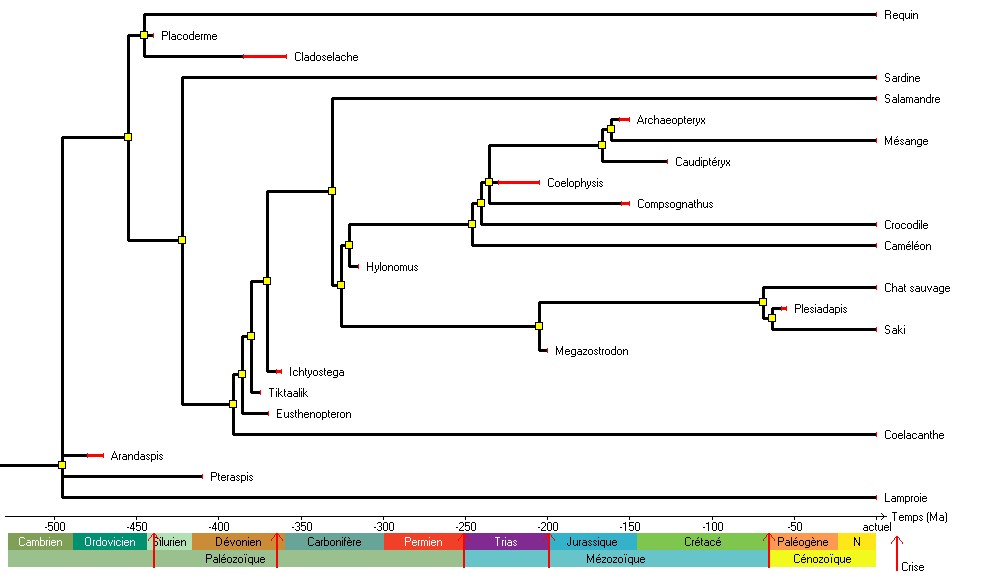
Figure 7. L'arbre des Vertébrés et une estimation des durées d'évolution (à partir des seuls fossiles de la collection)
6 - Phylogène permet d’utiliser la comparaison des séquences de gènes ou de protéines homologues pour établir des relations de parenté, avec les limites liées à la méthode phénétique utilisée. Mais on peut ne pas se limiter à la différence globale entre les séquences mais faire saisir qu’il y a eu, au niveau moléculaire ,une évolution dans les diverses branches. C’est l’intérêt de la fonction qui permet de chiffrer les changements sur chacune des branches de l’arbre. Les données relatives aux primates Cercopithécoïdes peuvent être ainsi utilisées pour visualiser une évolution très buissonnante dans cette lignée.
D’une façon plus qualitative, il est possible d’essayer de reconstituer dans des cas simples la séquence ancestrale d’un gène à partir des séquences des taxons actuels et d’indiquer la nature des innovations sur les différentes branches. D’une façon plus générale, il est bon de sensibiliser à l’idée qu’il y a eu des innovations génétiques originales dans chaque lignée. Un exemple intéressant est celui des Colobes, primates au régime alimentaire spécialisé (feuilles qu’ils digèrent grâce à des bactéries symbiotiques, un peu comme les ruminants, et ils se nourrissent finalement des produits de la digestion de ces bactéries). En rapport avec ce régime « bactérien » ils ont une grande richesse en ribonucléases pancréatiques, liée à une duplication du gène qui code pour la ribonucléase suivie de mutations ponctuelles qui rendent fonctionnelles ces ribonucléases dans des conditions de pH moins basique. Cette duplication ne se retrouve pas chez les autres primates.
III – Les erreurs relatives à la lecture des relations de parenté sur un arbre
Une mauvaise lecture des relations de parenté est due en partie aux conceptions sur le degré d’évolution des espèces et apparaît en particulier lorsque l’espèce jugée primitive est située à gauche sur l’arbre et l’espèce jugée la plus évoluée tout à fait à droite, comme sur l’arbre présenté précédemment. Elle résulte parfois aussi de la confusion entre similitude apparente des organismes et parenté. Deux grands types d’erreur sont signalés par les chercheurs américains :
A – Premier type d'erreurs
La proximité des taxons à l’extrémité des branches de l’arbre fournit des indications sur la parenté
Deux espèces, proches sur l'arbre, sont perçues comme plus étroitement apparentées que deux espèces éloignées. Leur ressemblance morphologique renforce encore cette lecture erronée.
Ainsi, sur l’arbre (Figure 8), la Sardine peut sembler plus apparentée au Requin qu’au Lézard du fait de sa position à côté du Requin.
Leurs morphologies les rapprochent encore (idées d'une classification par ressemblance) Le poids de l'histoire augmente encore la difficulté: ce sont deux poissons (groupe invalidé par l'approche phylogénétique moderne)
Figure 8. Erreurs liées à la proximité
Il faut donc amener les élèves à modifier leurs critères et à raisonner à partir des ancêtres communs qu’ils partagent ou non. Cela implique de lire le temps relatif sur l’arbre, ce qui peut être visualisé par des flèches sur les branches (fFgure 9) . Ainsi une chronologie relative des ancêtres hypothétiques est faite.
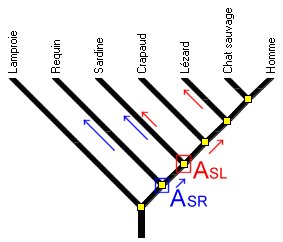
Figure 9a. Des flèches renforcent l'idée de temps et donc d'histoire évolutive. Le dernier ancêtre commun de la Sardine et du Requin (ASR) est plus ancien que le dernier ancêtre commun de la Sardine et du Lézard (ASL).
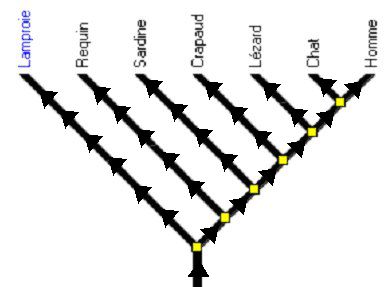
Figure 9b. Des flèches pourraient être dessinées manuellement sur les branches.
Type d'exercices proposé :
Pour entrainer les élèves à cibler uniquement le raisonnement sur les ancêtres, un bon moyen est de leur proposer l’analyse d’arbres d’aspect différent mais indiquant les mêmes relations de parenté.
B – Second type d'erreurs
Le nombre de nœuds est un indicateur des relations de parenté
Considérons l’arbre présenté à la figure 10 ( Extrait de la sous collection des Amniotes de « Phylogène » ).
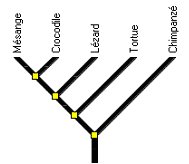
Figure 10 : La place du Chimpanzé.
Quand on demande d’indiquer qui de la Tortue ou de la Mésange est le plus étroitement apparentée au Chimpanzé, de nombreux élèves comptent le nombre de nœuds séparant la Mésange du Chimpanzé et le nombre de nœuds séparant la Tortue du Chimpanzé. Ils trouvent 4 nœuds entre la Mésange et le Chimpanzé et seulement 2 nœuds entre la Tortue et le Chimpanzé ce qui les conduit à dire que la Tortue est plus étroitement apparentée au Chimpanzé que ne l’est la Mésange.
Cela signifie qu’ils ne ciblent pas sur les ancêtres communs partagés par les deux espèces avec le Chimpanzé, ne repèrent pas donc pas qu’elles ont le même ancêtre commun avec le Chimpanzé et donc sont également apparentés au Chimpanzé.
Type d'exercices proposés :
Pour les aider à corriger leur pratique, on peut leur suggérer d’imaginer l’arbre si on supprimait le crocodile, puis le lézard.
On peut aussi partir du principe d'établissement des phylogénies: rechercher l'état de caractère partagé de manière exclusive par la tortue et le chimpanzé (groupe 1) puis par la mésange et le chimpanzé (groupe 2). On ne trouvera aucun état exclusivement partagé par un groupe. La fonction "Exploration des arbres" permet de rechercher facilement les états dérivés partagés exclusivement.
On peut noter à ce sujet que les questions posées au bac S visent toujours à démontrer qu’une espèce A est plus apparentée à B qu’à C mais jamais que l'espèce C est également apparentée à A et B, (figure 11) ce qui est autant révélateur de l’aptitude à lire correctement un arbre phylogénétique.
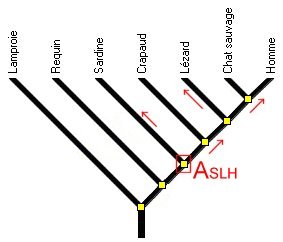
Figure 11. Le degré de parenté : la sardine est également apparentée au Lézard et à l'Homme. Le dernier ancêtre commun est le même dans les deux cas.
IV - Les erreurs dans la reconstitution des caractères des ancêtres communs hypothétiques
Ces erreurs sont liées aux idées fausses sur l’existence d’espèces ancestrales et d’espèces primitives et donc sur l’idée que des deux groupes issus d’un nœud, l’un puisse être l’ancêtre d’un autre et donc donner une image de l’ancêtre.
Par exemple, avec la collection sur les Cétacés, on établit que parmi les divers clades de mammifères actuels, c’est avec les Hippopotames que les Cétacés sont le plus étroitement apparentés, donc partagent un ancêtre commun exclusif. La tendance est forte alors d’imaginer que cet ancêtre commun est proche voisin des Hippopotames actuels, ce qui sous entend qu’il n’y a eu que peu d’évolution dans la lignée des Hippopotamidés. Cela est évidemment faux comme le suggèrent d’ailleurs les plus anciens fossiles de Cétacés. En revanche ceux-ci possèdent en commun avec les Hippopotames actuels des états dérivés au sein des Mammifères, comme l’astragale en double poulie que devait posséder l’ancêtre commun.
Cela étant, il est évident que l’évolution anatomique dans les deux lignées issues d’un nœud peut avoir été faible dans une lignée et beaucoup plus importante dans l’autre. Il est sans doute vrai que l’ancêtre commun aux crocodiles et oiseaux actuels devait davantage ressembler aux crocodiles qu’aux oiseaux sans être un crocodile pour cela ! Et bien sûr c’est toute la difficulté à ne pas décrire l’ancêtre commun à l’homme et au chimpanzé comme s’il était un chimpanzé, en l’absence de documents sur l’évolution dans la lignée des Paninés.
En guise de conclusion
L’interprétation d’arbres phylogénétiques présuppose l’idée d’évolution mais c’est aussi un passage nécessaire pour aboutir à une meilleure compréhension de ce qu’elle est et de ce qu’elle n’est pas, pour contribuer à faire disparaître les représentations initiales fausses. Cela exige l’analyse d’exemples choisis en fonction d’objectifs précis et va donc au-delà de la capacité à répondre correctement aux questions de bac du type II1. (Ces questions dans leur ensemble testent l’aptitude à indiquer les parentés à partir d’un arbre mais pas la compréhension d’ensemble de l’évolution).
Cette compréhension de l’évolution, cette structuration de l’histoire des êtres vivants est une base indispensable pour aborder les mécanismes de l’évolution (process). En fin de compte ce qu’il s’agit de tenter d’expliquer, ce sont les mécanismes qui ont contribué à cette histoire. La dissociation actuelle dans les programmes de terminale S de l’établissement des phylogénies (première partie du programme) et des mécanismes de l’évolution (noyés dans l’étude du brassage génétique assuré par les mécanismes de la reproduction sexuée) est regrettable.
A vrai dire, l’étude des mécanismes de l’évolution est aussi utile pour une bonne compréhension de ce qu’est un arbre car en fin de compte quelles idées doit avoir un élève de ce qui se passe le long des branches de l’arbre, des mécanismes en œuvre aux nœuds qui aboutissent à la genèse de deux lignées, et finalement de ce qu’est un ancêtre commun ? L’introduction du mode de pensée populationnel est nécessaire et faire saisir qu’un nœud correspond à un évènement de spéciation, même si cela n’est que rarement indiqué dans les textes consacrés à l’établissement des phylogénies (l’abandon du problème de la spéciation est sans aucun doute une lacune du programme de TS ).
Jusqu’ici, Phylogène était un logiciel ciblé sur la construction d’arbres phylogénétiques à partir de l’observation, la comparaison des organismes, etc. Il y avait une certaine limite pratique à son utilisation dans la mesure où à l’examen, il est exclu de demander aux lycéens de construire un arbre phylogénétique à partir d’une matrice de caractères. Néanmoins, apprendre à construire un arbre à travers des actions dont on comprend la signification, facilite ensuite l’interprétation d’arbres fournis à l’examen. En outre, son utilisation fournit une aide pour répondre à une question de type I comme celle posée en juin 2009 « Après avoir exposé les principes permettant d’établir les relations de parenté … »
La possibilité de faire des exercices à partir d'arbres déjà construits a été pensée dans cet esprit. Ces arbres restent dynamiques : on peut y rechercher les états de caractères et les portraits robots des ancêtres. Suivant les choix faits par le professeur à l'enregistrement, l'arbre peut être ou non éditable. Un arbre éditable est un arbre dont la descendance des ancêtres peut être modifiée. Permuter l'ordre des branches reste toujours possible.
1 Nous nous plaçons ici dans le cadre de la macroévolution (le seul cadre pertinent pour Phylogène).
2 S.J. Gould Comme les huit doigts de la main – Edition du Seuil (1993) p302
Bibliographie :
D. Baum, S. Smith, S. Donovan (2005) The tree thinking challenge. Science, 310, 979-980
M. Crisp, L. Cook ( 2005) Do early branching lineages signify ancestral traits? Trends in Ecology and Evolution, 20,122-128
S. Nee (2005) The great chain of being, Nature, 435, 429
K. Omland, L. Cook, M. Crisp (2008) Tree thinking for all biology : the problem with reading phylogenetics as ladders of progress. Bioessays, 30, 854-867
T. Ryan Gregory (2008) Understanding evolutionary trees. Evolution: Education and outreach, volume 1(2) , 121-137
B. D. Boucher
Enseigner l'évolution de la lignée humaine, le "retour des vieux démons" ?