
Structures et circuits de l'état de veille
Les structures nerveuses et les circuits impliqués dans l'état de veille
Pascal Convers, Françoise Jauzein, INRP, ERTé ACCES, 2005
L'éveil correspond à la sortie du sommeil. Ainsi l'état de veille peut être défini comme l'état correspondant à la période allant du réveil à l'endormissement. L'éveil au niveau cortical correspond à un EEG (électroencéphalogramme) caractérisé par des ondes d'amplitudes faibles (50.1O -6 volts) et de fréquence élevée (20 hertz): on les appelle les ondes alpha.
NB: on enregistre également les mouvements occulaire et le tonus musculaire pour caractériser les différentes étapes du sommeil.
Figure 1. Représentations de quelques enregistrements effectués lors de l'endormissement
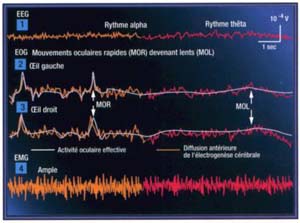
L'activité corticale enregistrée pendant la veille est dite désynchronisée: les différents neurones fonctionnent de façon indépendante. Elle s'oppose à la synchronisation observée pendant le sommeil . L'éveil correspond donc à la désynchronisation corticale.
Le cortex cérébral ne possède pas de mécanismes intrinsèques propres à son activation (=désynchronisation). En effet une préparation de cerveau isolé se caractérise par un EEG avec des ondes lentes permanentes de haute amplitude ressemblant à celles du sommeil profond (cf. ci-dessous)
Figure 2. Schéma de l'expérience réalisée Par Bremer en 1935
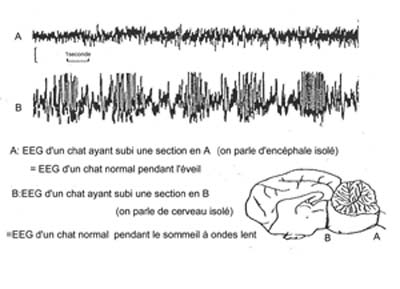
Cette expérience montre que des structures sous-corticales sont responsables de l'activation corticale (=désynchronisation). Ce sont ces structures qui seront décrites dans la suite de cet article. Les principales structures seront classées selon l'ordre de leurs découvertes. On trouvera ensuite un bilan de l'état actuel des connaissances concernant les réseaux de neurones impliqués dans l'éveil.
L' hypothalamus postérieur
L'observation historique de Von Ecomo
La grippe espagnole (=encéphalite épidémique qui a fait 25 millions de morts dans le monde) est survenue à Vienne en 1920. On distinguait deux types de malades: les agités qui criaient tout le temps et ne dormaient pas et ceux qui étaient dans le coma (impossible de les réveiller). Beaucoup de ces patients moururent. Constantin Von Economo (neurologue viennois ) étudia le cerveau des deux groupes et exposa ses résultats en 1928. Il démontra que les malades qui ne dormaient pas présentaient une lésion (due au virus de l'encéphalite) dans la partie antérieure de l'hypothalamus (région préoptique). La lésion avait détruit un centre du sommeil ("Schlafzentrum"). Ceux qui restaient dans le coma présentaient une lésion de l'hypothalamus postérieur ou de la région haute du mésencéphale. Von Economo postula l'existence d'un centre de l'éveil (=Wachzentrum) situé dans l'hypothalamus postérieur (d'après M Jouvet, Pourquoi rêvons-nous? pourquoi dormons-nous?, Editions Odile Jacob, 2000,120 pages).
Figure 3. Localisation de l' hypothalamus antérieur et postérieur
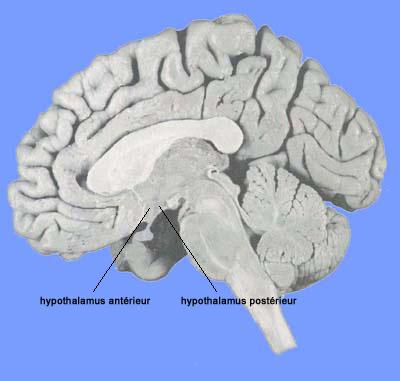
Quelques observations complémentaires
-
Les patients ayant une lésion inflammatoire de l'hypothalamus postérieur sont atteints d'hypersomnie
-
Des lésions expérimentales de cette zone sur différentes espèces (singe, rat, chat) se traduisent également par des hypersomnies
La formation réticulée mésencéphalique
En 1949 Giuseppe Moruzzi et Horace Magoun déclenchent un coma chez le chat en lésant la formation réticulée (=zone présentant un aspect réticulé et située dans le tronc cérébral).
Les mêmes scientifiques stimulent électriquement un groupe de noyaux cholinergiques situés à la jonction du pont et du bulbe et produisent ainsi un état de vigilance et d'éveil. En effet, leur stimulation provoque la "désynchronisation" de l'EEG (= l'activité EEG passe d'un tracé d'ondes synchronisées de haute amplitude à des ondes désynchronisées, de plus haute fréquence et de plus faible amplitude).
Cette zone est donc appelée système réticulaire activateur (= formation réticulée mésencéphalique) et Moruzzi et Magoun proposent la théorie réticulaire. Selon celle-ci l'activation corticale est sous la dépendance du système réticulaire par l'intermédiaire de 2 systèmes ascendants, soit le réticulo-thalamo-cortical (en rouge), soit directement, le réticulo-cortical (en bleu).
Figure 4. Schéma de la thorie réticulaire
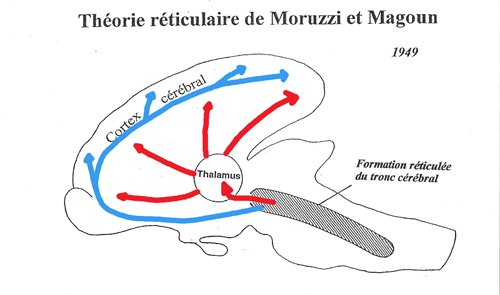
Cependant des lésions de la formation réticulée et de l'hypothalamus postérieur ne perturbent pas durablement l'éveil: d'autres structures prennent donc le relai des structures détruites. De plus la lésion sélective des corps cellulaires des neurones (possible dès 1983 grâce à l'injection in situ d'acide kainique ou iboténique qui entraîne une dépolarisation intense des corps cellulaires et provoque leur mort mais permet de conserver les axones intacts) de la formation réticulée mésencéphalique n'entraîne aucun trouble de l'éveil comportemental, ni aucune altération de l'activation corticale. Ainsi les troubles de l'éveil obtenus précédemment résultaient sans doute de lésion des prolongemnents des neurones localisés dans les autres systèmes de l'éveil.
A l'heure actuelle de nombreuses expériences démontrent que les systèmes d'éveil sont disposés en réseaux, c'est-à-dire que l'excitation pharmacologique de l'un est suivie par l'activation de tous. De même, cette organisation en réseau redondante explique pourquoi l'inactivation d'un seul système est suivie après quelques jours par une récupération quasi complète de l'éveil. Il est donc difficile de comprendre comment fonctionne la plupart de ces systèmes organisés en réseaux. Il existe cependant, à certains "noeuds" de ces réseaux, des endroits dont l'inactivation peut inhiber tout le réseau. L'organisation des réseaux exécutifs peut se résumer comme cela est décrit ci-dessous.
Les réseaux exécutifs de l'éveil
Les réseaux encéphaliques
-
le système de l'hypothalamus postérieur
-
Les neurones à histamine.
lls sont localisés dans le noyau tubéromammillaire et la région adjacente de l'hypothalamus postérieur.
Figure 5. Noyau tubéromammillaire et projections histaminiques
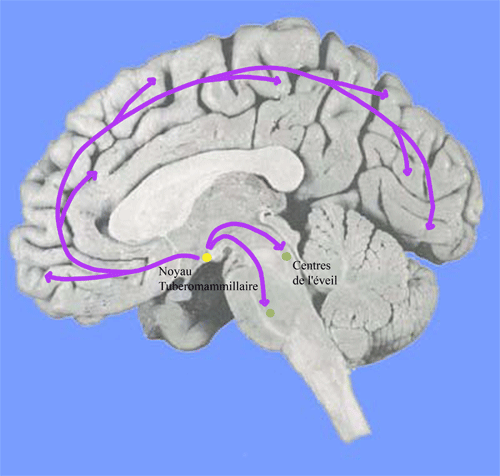
Ils augmentent leur activité pendant l'éveil. Ils ont des projections diffuses dans la majorité des régions cérébrales ( en particulier au niveau des autres systèmes de l'éveil) et ont une action activatrice. Ils contribuent ainsi à la genèse d'une activation corticale généralisée et au maintien de l'éveil. L'inactivation des neurones à histamine entraîne le sommeil lent. Le blocage des récepteurs H1 à l'histamine entraîne une diminution de l'éveil et une augmentation de sommeil lent (remarque: les médicaments antihistaminiques ont un effet sédatif).
Pour plus de détail consulter l' article structures et mécanismes responsables du cycle veille sommeil disponible sur le site le sommeil, les rêves et l' éveil de l'univerité Lyon 1.
-
Les neurones à orexine.
Leurs corps cellulaires sont situés dans la partie dorso-latérale de l'hypothalamus postérieur c'est à dire dans une position plus dorsale que les neurones à histamine. Les neurones à orexine sont actifs pendant l'éveil; leur activité diminue pendant le sommeil lent et ils sont inactifs pendant le sommeil paradoxal. L'injection d'orexine stimule l'éveil (soit directement soit indirectement par stimulation de la prise alimentaire).
Les neurones à orexine ont été découverts il y a 5 ans. L'orexine a la même structure que la sécrétine et elle a été mise en évidence dans l'hypothalamus d'où son nom d'hypocrétine. Elle a été appelé Orexine (= qui stimule la prise alimentaire) par un groupe travaillant sur la prise alimentaire. Elle aurait un rôle dans la narcolepsie puisque son absence déclenche la narcolepsie (mais on ne sait pas par quel mécanisme).
-
-
Le système du télencéphale basal
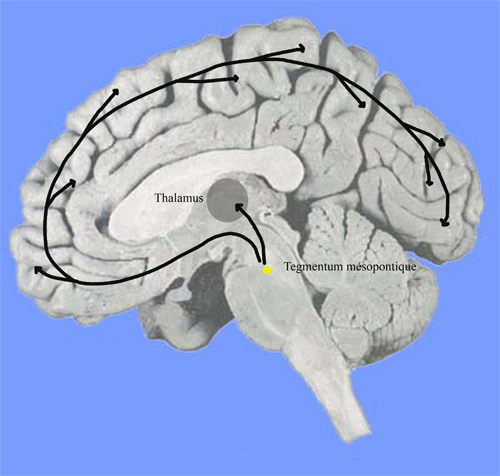
Le maillon principal de ce système est constitué de neurones synthétisant l'Acétylcholine et/ou le GABA. Ces neurones envoient des projections à tout le cortex et aux noyaux thalamiques. Leur stimulation est éveillante mais leur lésion neurotoxique n'entraîne qu'une diminution très transitoire de l'éveil. Ils sont situés dans le tegmentum mésopontique à la jonction du pont et du mésencéphale.Ils se projettent sur les neurones thalamocorticaux. Un grand nombre de ces neurones présente une fréquence de décharge élevée durant la veille et le sommeil paradoxal. A l'inverse, ils sont au repos pendant le sommeil lent.
Figure 6. Tegmentum mésopontique et projections cholinergiques ou GABAergiques
-
Le système thalamique
Les neurones thalamiques intralaminaires se projettent à l'ensemble du cortex. Un de leurs neurotransmetteurs est un acide aminé excitateur (aspartate/glutamate).
Les systèmes du tronc cérébral
-
La formation réticulée mésencéphalique (=FRM)
Elle contient des neurones qui sécrètent de acides aminés excitateurs comme l'aspartate et le glutamate (neurones Asp/Glu). Ils se projettent massivement sur les noyaux thalamiques. La stimulation de ces neurones déclenche un éveil prolongé. Avec les neurones cholinergiques mésopontins (voir paragraphe suivant), la FRM forme la voie réticulo-thalamo-corticale. Ainsi, la formation réticulée (l'ancien système d'éveil) est devenue une partie du réseau de l'éveil.
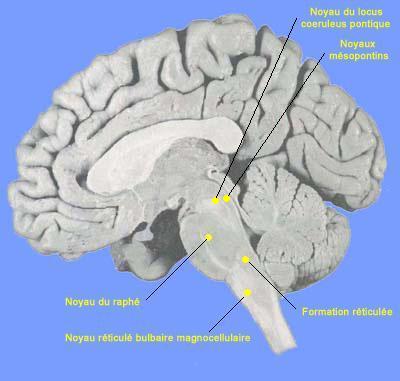
Figure 7. Localisation des noyaux du système du tronc cérébral
-
Les noyaux mésopontins cholinergiques
Les neurones de ces noyaux se projettent en partie sur le thalamus. L'acétylcholine exerce une double action : une action muscarinique inhibitrice par hyperpolarisation des neurones réticulaires (appartenant au système du sommeil) et une action nicotinique activatrice par dépolarisation des neurones thalamocorticaux et corticaux.
-
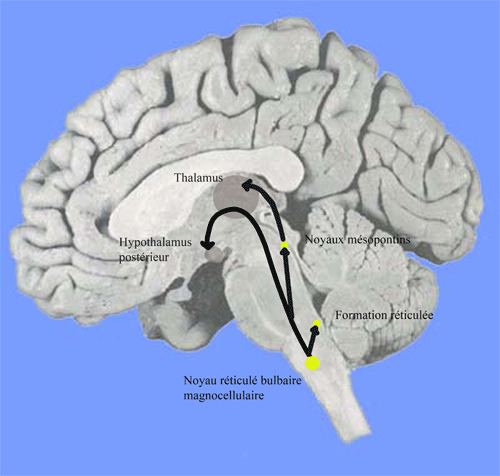
Le noyau réticulé bulbaire magnocellulaire (MC)
Les neurones sont cholinergiques ou Asp/Glu-ergiques. Ils se projettent sur la formation réticulée mésencéphalique, l'hypothalamus postérieur et les groupes cholinergiques mésopontins et du télencéphale basal. Cet ensemble forme la voie réticulo-hypothalamo-corticale. La stimulation du noyau magno-cellulaire provoque un éveil intense et prolongé
Figure 8. Système cholinergique de l'éveil
-
Les neurones sérotoninergiques des noyaux du raphé
Le raphé antérieur contient des neurones contenant la 5-hydroxytryptamine (5HT) ou sérotonine. Ils se projettent vers l'hypothalamus et le cortex. Ces neurones sont actifs pendant l'éveil. Leur stimulation globale est éveillante et, contrairement aux autres groupes aminergiques, leur lésion, non seulement n'entraîne pas de somnolence, même passagère, mais provoque une insomnie prolongée de plusieurs jours. On peut démontrer le rôle éveillant du système à sérotonine, soit en enregistrant les corps cellulaires pendant le cycle éveil-sommeil, soit en inactivant les corps cellulaires par injection in situ d'agonistes des récepteurs H3.
Figure 9. Système sérotoninergique de l'éveil
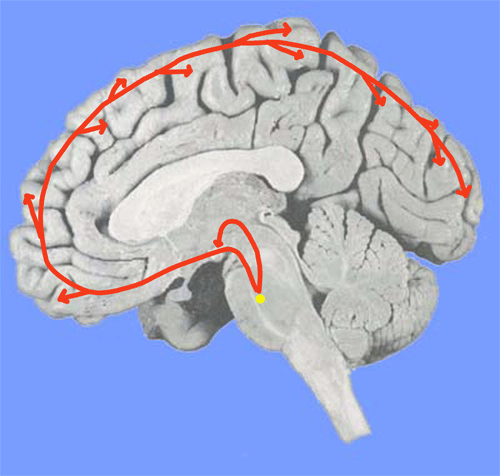
-
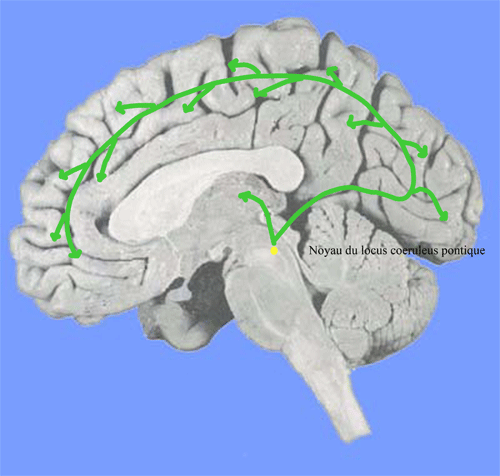
Le noyau du locus coeruleus pontique
Il contient des neurones noradrénergiques (NA) qui envoient des projections directes vers le cortex, le thalamus et l'hippocampe. Leur activité unitaire augmente pendant l'éveil et diminue pendant le sommeil. L'inhibition de la synthèse de la noradrénaline (NA) entraîne une synchronisation corticale typique du sommeil lent. Ces neurones reçoivent de nombreuses afférences activatrices et inhibitrices.
Figure 10. Système Noradrénergique de l'éveil
Bilan
La régulation de l'éveil est un phénomène complexe mettant en jeu des structures multiples et redondantes. Aucune des structures décrites, prises isolément, n'est indispensable à l'activation corticale. Ainsi les différents systèmes impliqués sont connus mais il reste à determiner comment le fonctionnement en réseau de ces différents systèmes permet la régulation de l'éveil.