
Le développement des voix visuelles
LE DEVELOPPEMENT DES VOIES VISUELLES CHEZ L’ ENFANT ,
CHAPITRE 1 : DEVELOPPEMENT DES VOIES VISUELLES ANTERIEURES
PENDANT LA PERIODE PRENATALE
Introduction
I - Le développement du globe oculaire :
II - Formation du nerf optique et de la papille :
III - Formation du cristallin :
CHAPITRE 2 : LE DEVELOPPEMENT DE LA VISION CHEZ L’ENFANT
I - Les voies sensori-gnosiques : la vision
1. Le récepteur : la rétine
b) Développement
b) Le développement
3. Le cortex visuel
b) Le développement
II – Le décodage du sens : le niveau gnosique
CONCLUSION
CHAPITRE 3 : ACUITE VISUELLE CHEZ UN ENFANT D’AGE PREVERBAL
I- Méthodes comportementales subjectives de mesure du pouvoir séparateur
1.Technique du regard préférentiel ou « Bébé vision »
c) Résultats de la technique du regard préférentiel
2 . La poursuite visuelle d'une mire structurée en mouvement
II – Méthodes comportementales objectives du pouvoir séparateur
Le nystagmus optocinétique (NOC)
a) Définition :
b) Principe :
c) Résultat :
III – Acuite visuelle et maturation par potentiels évoqués visuels
1. Par extrapolation
2. Les facteurs de variation de l'acuité visuelle par PEV chez l'enfant
IV – Comparaison des différentes méthodes de mesure du pouvoir séparateur chez le jeune enfant
CONCLUSION
CHAPITRE 1 : DEVELOPPEMENT DES VOIES VISUELLES ANTERIEURES
Introduction
La vision permet à un organisme animal de s’informer sur son environnement. Chez les vertébrés, cette capacité est rendue possible par l’existence d’organes sensoriels : les yeux et en particulier la rétine.
La rétine est une excroissance du cerveau, qui se forme pendant le développement embryonnaire à partir du diencéphale. Elle est composée de divers types cellulaires, en particulier ceux capables de détecter certains stimulus lumineux : les cônes et les bâtonnets. Toutefois, l’œil en lui-même n’est pas capable d’interpréter les informations ainsi perçues. Cette interprétation est réalisée par l’encéphale de l’animal. Ainsi, les informations captées par les cellules de la rétine sont véhiculées, sous forme de message nerveux, depuis l’œil jusqu’au cerveau, par le nerf optique. On parle pour cela des voies visuelles primaires. Chez les mammifères, les nerfs optiques véhiculent une information nerveuse qui, après relais au niveau du thalamus, aboutit au cortex occipital. Après les premières interprétations (forme du stimulus, couleur, mouvement, etc..) des mécanismes plus complexes (pouvant mettre en jeu d’autres zones de l’encéphale) aboutissent à la représentation consciente du stimulus par l’animal, et par voie de conséquence à une modification appropriée de son comportement.
I - Le développement du globe oculaire
Le développement des voies visuelles antérieures met en jeu le neuro-ectoderme, l'ectoderme de surface, la crête neurale et le mésoderme à un moindre degré.
- 3ème semaine : premières ébauches du globe oculaire, les fossettes optiques, sous la forme de 2 dépressions symétriques sur les parois latérales de la gouttière neurale. A la fermeture du tube neural, elles s'évaginent et deviennent les vésicules optiques, unies au tube neural par le pédicule optique ou tige optique. La convexité des vésicules optiques se retrouvent ainsi au contact de l'ectoderme.
- 4ème-6ème semaines : formation de la cupule optique, par aplatissement et invagination de la vésicule optique. Les bords inférieurs de la cupule sont séparés par une fissure, la fissure ftale ou fente choroïdienne ou fente ftale ou fente embryonnaire, qui s'étend sur le pédicule optique.
La cupule comprend ainsi 2 couches : une couche externe qui deviendra l'épithélium pigmenté, une couche interne qui donnera la rétine entre autres ; ces 2 couches sont séparées par un espace virtuel, qui s'il persiste au cours du développement est responsable d'un décollement de rétine congénital ou plus exactement d'une absence d'attachement des feuillets.
En même temps que la formation de la cupule optique, l'ébauche du cristallin s'individualise à partir de l'ectoderme, en regard de la vésicule puis de la cupule optique. La placode du cristallin se dessine à la 4ème semaine, puis se déprime pour former la fossette cristallinienne ou fovea lentis, puis elle s'élargit pour former la vésicule cristallinienne, qui se sépare de l'ectoderme au début de la 5ème semaine pour s'incorporer dans l'ouverture de la cupule optique.
Le mésenchyme qui entoure la vésicule optique (cellules de la crête neurale) va pénétrer à l'intérieur de la cupule par la fente ftale et formera le vitré primitif.
Les vaisseaux embryonnaires apparaissent : à la fin de la 4ème semaine 2 plexus s'isolent de l'artère ophtalmique branche de la carotide interne : -l'artère hyaloide se ramifie dans le vitré et se termine à la partie postérieure du cristallin pour former la tunique vasculaire du cristallin; -l'artère marginale se divise en 2 branches et s'étalent à la surface de la cupule optique.
- 5ème-6ème semaines : fermeture de la fente ftale alors que la cupule optique reste ouverte en avant.
- 6ème semaine : formation du vitré secondaire définitif. Le vitré primitif est une structure collagénique, vascularisée. Le vitré secondaire devient avasculaire, en particulier par synthèse de l'acide hyaluronique. Les vaisseaux hyaloïdiens s'atrophient avec formation du canal de Cloquet qui contient les restes du vitré primitif et de l'artère hyaloïde.
II - Formation du nerf optique et de la papille :
- Le nerf optique se forme au cours des deux premiers mois : la fermeture de la fente ftale débute au milieu du pédicule optique et se poursuit en avant et en arrière par fusion des feuillets externe et interne. A la 5ème semaine les axones nés des cellules ganglionnaires du feuillet interne de la cupule optique se dirige vers la future papille et isolent une saillie appelée la papille épithéliale de Bergmeister (de nature gliale). Les axones atteignent le chiasma au 45ème jour.
- Les gaines méningées sont visibles plus tard : la pie-mère au 4ème mois, la dure-mère au 5ème mois, l'arachnoïde au 6-7ème mois.
- La papille de Bergmeister se renfle au 4ème mois puis régresse au 5ème mois. L'importance de l'atrophie de la papille primitive règlera la profondeur de l'excavation physiologique de la papille.
III - Formation du cristallin :
- La vésicule cristallinienne formée, les cellules de la paroi antérieure diminuent de hauteur, alors que celles de la paroi postérieure s'allongent et formes les fibres primitives. A la 7ème semaine la cavité de la vésicule cristallinienne a disparu et le cristallin est un noyau plein. A la 8ème semaine l'épithélium postérieur transformé en fibres a disparu et la croissance du cristallin est assurée par les cellules situées à l'équateur (jonction des deux épithélium) connues sous le nom de vortex.
- La chambre antérieure est formée au 5ème mois.
CHAPITRE 2 : LE DEVELOPPEMENT DE LA VISION CHEZ L’ENFANT
Introduction :
Le développement du système visuel chez l’enfant est à connaître pour comprendre la physiopathogénie de l’amblyopie et du strabisme, et pour analyser l’examen de dépistage de la fonction visuelle obligatoire chez l’enfant au 8e mois.
Le système visuel de l’enfant n’est pas mature à la naissance. Une maturation post-natale s’observe au niveau rétinien, au niveau des voies visuelles et au niveau cortical et sous-cortical.
I - Les voies sensiro-gnosiques : la vision
Il s'agit des voies centripètes, afférentes, voies sensorielles qui vont conduire à l'analyse et au
1. Le récepteur : la rétine
a) Structure
A la naissance, la rétine périphérique est identique à celle de l'adulte. Par contre la rétine maculaire est profondément immature. Toutes les couches cellulaires y sont, mais de faible épaisseur. Les photorécepteurs sont formés d'un article interne et d'un article externe encore très court et ramassé. Toutes ces particularités apparaissent dès la région nasale adjacente à la fovéa et atteignent leur maximum au centre de cette fovéa.
Il est classique de penser que l'immaturité constituée par l'écartement des articles externes et leur faible contenu en pigment contribuent à la mauvaise acuité visuelle du nouveau-né dont la vision est davantage extrafovéolaire. Cependant, le nouveau-né est capable de fixation centrale quand il est placé dans de bonnes conditions.
b) Développement
Une rétine d'un nourrisson de 11 mois présente une morphologie de type adulte, la rétine périphérique est plus rapidement fonctionnelle que la zone centrale.
La surface totale de la rétine (épithélium pigmentaire) croît rapidement durant les derniers mois de gestation, puis plus graduellement lors des deux premières années post-natales à la fin desquelles, elle atteint sa taille adulte. Sa surface est alors la même que chez un adulte soit de 944 mm2.
Le développement de la structure maculaire est long et ne se rapproche du stade adulte que vers 45 mois.
- A 22 semaines de gestation, la zone fovéolaire est identifiable par la présence d'une fine couche de cellules ganglionnaires et d'une couche de photorécepteurs contenant uniquement des cônes. La fovéa est très immature à la naissance (fig 2). Sa maturation morphologique est marquée tout d'abord par la migration périphérique des couches rétiniennes centrales, formant ainsi la dépression fovéale qui continue de s'approfondir jusqu'à 15 mois. Dans un deuxième temps, nous assistons à un allongement et à une migration centrale des articles externes cônes.
- A la naissance, la zone sans bâtonnets ou fovéola a environ 1000 µm de diamètre, elle devient progressivement plus étroite pour atteindre la dimension adulte de l'ordre de 400 à 700 micron vers 45 mois. L'aire maculaire, dont les limites sont difficiles à déterminer, correspond à une surface de 1,7 mm2 à 3 mm2 soit un champ de 3 à 10 degrés. Parallèlement, la concentration des cônes augmente, 18 cônes / 100 µm à 1 semaine à 42 cônes / 100 µm chez un adulte.
Cependant bien que l'on considère la macula presque mature à 45 mois, on relève que la densité centrale des cônes et la longueur de leur segment externe, sont, à cette période à la moitié de leur valeur adulte. Ces éléments anatomiques contribuent largement à expliquer que l'acuité soit encore plus faible que chez l'adulte.
Les axones des cellules ganglionnaires de la rétine vont converger en direction de la papille puis former le nerf optique (dont la myélinisation s'achève totalement à la fin de la deuxième année). La papille du nouveau-né paraît pâle et devient rosée vers le 4 ème mois.
Les processus photochimiques rétiniens sont fonctionnels très rapidement. Des études électrorétinographiques sur la sensibilité scotopique et la régénération de la rhodopsine montrent que vers 10 semaines les amplitudes des ondes b scotopiques sont similaires à celles trouvés chez l'adulte (FULTON 1988).
La myélinisation des fibres ganglionnaires qui constituent le nerf optique, débute entre le 6ème et le 8ème mois de vie foetale et cette myélinisation se poursuit jusqu'à l'âge de 2 ans.
Figure 1 : sections verticales d'une rétine humaine d'un nouveau-né de 5 jours. Zone A : centre de la fovéa . Zone B 900µm en nasal du centre de la fovéa soit approximativement 5°. Zone C 1800µm en nasal du centre de la fovéa soit approximativement 10° . Un cône est souligné sur chaque photographie (CANDY 1998)

2. Le corps genouillé latéral (cGLd)
a) structure
Par des méthodes histologiques, la structure du cGLd est bien connue chez le singe et chez
l'homme. On retrouve les 6 couches, 2 magnocellulaires ventrales à grande cellules (1 et 2), 4
parvocellulaires dorsales à petites cellules (3, 4, 5, 6). On retrouve également la ségrégation oeil droit, oeil gauche). Entre ces deux ensembles de couches qui constituent les voies M et P, on décrit une multitude petites cellules situées entre les couches M et P, qui constituent la voie Koniocellulaire, ou voie K, qui aurait un rôle dans le maintien de l'inhibition entre les canaux de transmission des types de signaux (VITAL-DURAND 1999).
b) Développement
Chez l'homme, la lamination du cGLd est identifiable dés la 24ème semaine de gestation .Les différents types de neurones sont repérables vers la 35ème semaine d'âge gestationnel .
A la naissance le cGLd est déjà organisé en couches (6 couches).
Mais à la naissance ces cellules présentent des critères d'immaturité. Il existe de nombreuses épines sur le corps neuronal et sur les dendrites des cellules bipolaires et multipolaires. Le nombre d'épines atteint un maximum à l'âge de 4 mois puis diminue jusqu'à l'âge de 9 mois, les corps cellulaires et les dendrites arrivent alors à leur diamètre définitif. Le cGLd atteint sa taille adulte à 6 mois. Chez le singe, les neurones de cGLd sont formés entre le 36ème et le 43ème jour embryonnaire, puis pendant les deux premiers mois va se produire une maturation rapide avec une diminution des temps de latence, une diminution de la taille des champs récepteurs, une augmentation du pouvoir de résolution spatiale de la région fovéale (VITAL-DURAND 1985).
3. Le cortex visuel
a)Structure
Le cortex visuel primaire est une véritable carte rétinotopique composée de modules juxtaposés ou intriqués, de colonnes indépendantes. Les colonnes de dominance oculaire représentent des bandes de tissu cortical alternativement occupées par des afférences issues de l'oeil gauche ou de l'oeil droit. Ces bandes sont particulièrement marquées au niveau de la couche IV du cortex, couche qui reçoit les terminaisons des afférences du corps genouillé latéral, et où les neurones sont exclusivement activés par la stimulation de l'un ou l'autre oeil, mais sont encore visibles dans les couches supra ou infragranulaires où les neurones sont préférentiellement activés par l'un ou l'autre oeil.
Les colonnes d'orientation représentent des bandes fines de tissu cérébral à l'intérieur desquelles les neurones sont sélectifs par exemple à la même orientation du stimulus visuel.
Ces colonnes d'orientation traversent toutes les couches du cortex cérébral, à l'exception de la
couche IV où les neurones ne présentent pas de sélectivité à l'orientation. Ces colonnes d'orientation sont orthogonales aux colonnes de dominance oculaire, ce qui traduit leur indépendance. On parle alors d'hyper colonne .
Depuis quelques années, a été mis en évidence, grâce à l'analyse de la distribution dans le cortex visuel primaire d'une enzyme mitochondriale, la cytochrome oxydase un aspect important de l'architecture fonctionnelle du cortex strié (IMBERT 2000). Lorsque des coupes de cortex sont traitées pour révéler la cytochrome oxydase, on peut observer une coloration continue dans la couche IV et une coloration discontinue, par petits paquets ovoïdes périodiques appelés blobules, dans les couches superficielles et profondes. Sur des coupes tangentielles, passant dans le plan des couches II-III, les blobules de cytochrome oxydase apparaissent comme des colonnes vues selon leurs sections transverses, centrées sur les colonnes de dominance oculaire.
Cette différenciation cytochimique, désignée du terme anglais de " blobs " est fortement corrélée avec une ségrégation fonctionnelle nouvelle (fig. 3). En effet, les neurones enregistrés à l'intérieur des blobules sont pour la plupart non sélectifs à l'orientation : ils répondent de façon optimale aux basses fréquences spatiales, quelle que soit l'orientation du stimulus rectiligne employé ; en revanche, plus de la moitié d'entre elles sont sélectives à la longueur d'onde de la lumière utilisée.
Les cellules enregistrées en dehors des blobules sont au contraire sélectives à l'orientation, mais pas, pour la grande majorité d'entre elles, à la longueur d'onde .
Les cellules ganglionnaires issues de chaque rétine se projettent dans les couches magno-cellulaire 2 (2 m) et parvocellulaire 3 (3 p) du corps genouillé latéral (CGL) ipsilatéral et dans les couches magnocellulaire1 (1 m) et parvo-cellulaire 4 (4 p) du corps genouillé latéral controlatéral. Les axones thalamiques liés au système magno-cellulaire se projettent dans la couche IVC, ceux liés au système parvo-cellulaire dans la couche IVCb du cortex visuel primaire, indépendamment pour l’oeil droit et pour l’oeil gauche. Les colonnes d'orientation (CO) sont orthogonales aux colonnes de dominance oculaire (CDO). Modifiée d'après M. IMBERT, C. FONTA. Les Séminaires ophtalmologiques d'IPSEN, tome 11" Interfaces et interaction de la rétine ". Ed Y. Christen, M. Doly, M.-T. Droy-Lefaix, eds, © 2000, Irvinn, Paris, 35-47

Figure 2 Organisation de la voie rétino-thalamo-corticale chez le marmouset
Au total, le cortex visuel primaire peut être considéré, jusqu'à un certain point, comme un "séparateur d'attributs " : il permet en effet une séparation dans des territoires distincts du codage des orientations, de la couleur, de la forme et de la distance des objets de l'environnement spatial. A chacun des attributs codés correspondent un circuit et un domaine neuronal particulier : colonnes ou bandes, strates ou amas (blobules).
Relayée par le corps genouillé latéral (cGLd), l'information visuelle, codée par la rétine, arrive au niveau du cortex cérébral dans la partie la plus postérieure du lobe occipital. Les axones des neurones relais du cGLd se terminent presque exclusivement dans des couches déterminées (principalement la couche IV) de l'aire 17 de Brodmann (ou aire visuelle striée, ou V1). Dans le cortex primaire, les fibres afférentes thalamiques maintiennent l'ordre
topologique de la distribution rétinienne des cellules ganglionnaires d'origine, réalisant ainsi une carte précise du demi-champ de vision controlatéral. De là l'information est distribuée sur les 20 ou 30 autres aires corticales dans lesquelles les signaux circulent dans les deux sens, chaque aire spécialisée préparant les autres à traiter une partie du signal.
b) Développement
Il faut du temps pour qu'un cerveau parvienne à sa pleine maturité. Dans l'espèce humaine, près de 20 ans sont nécessaires pour que la myélinisation soit complète. La complexité du cerveau tient non seulement au fait qu'il est composé de très nombreux éléments, mais réside dans la richesse et la précisions des interconnexions entre ces très nombreux éléments : chaque cellule nerveuse reçoit ou est à l'origine de 5000 à 90 000 contacts fonctionnels à travers lesquelles les cellules nerveuse communiquent entre elles pour capter échanger, traiter, stocker et confronter des informations pertinentes qui assurent à l'organisme un comportement biologiquement adapté.
Dans la phase précoce du développement, les neurones, formés en nombre supérieur à celui qui constituera le stock adulte, doivent se déplacer sur de grandes distances. Pendant cette phase, les connexions réalisées sont relativement grossières et vont esquisser un cerveau seulement approximatif.
C'est au cours d'une seconde phase, plus tardive, que cette ébauche sera affinée, épurée, en particulier par des mécanismes d'élimination sélective (mort neuronale et apoptose), réduisant par mort cellulaire le nombre de neurones et de nombreuses connexions. Cette élimination massive commence avant la naissance. Elle continue après la naissance avec une perte de plus de 15% des neurones dans les premières semaines de la vie.
Chez tous les mammifères, le cortex visuel primaire est immature à la naissance. Des techniques autoradiographiques des voies nerveuses reliant entre elles les neurones des diverses structures visuelles - rétine, cGLd, cortex cérébral, associées à des techniques de traçage, antérograde et/ou rétrograde, ont permis de préciser le calendrier des étapes de mise en place des principales structures visuelles et de leurs interconnexions.
L'expérience visuelle précoce, que le jeune enfant éprouve, en particulier au cours de périodes dites critiques, régit le développement et le maintien des propriétés physiologiques et anatomiques caractéristiques de ce système chez l'adulte.
La plasticité du développement postnatal du système visuel dépend d’événements anatomiques, notamment la ségrégation en modules fonctionnels, qui sont sous la dépendance de cette expérience qui commande, en outre, l'acquisition des propriétés fonctionnelles spécifiques de ce réseau neuronal.
Il existe une période sensible, correspondant à la phase de développement au cours de laquelle le fonctionnement du système visuel est particulièrement dépendant de la qualité de l’expérience visuelle dont bénéficie le sujet.
La période sensible a été largement étudiée en privant les animaux de vision par tarsorapie. C’est ainsi que l’on peut montrer qu’une période de privation monoculaire d’expérience visuelle de quelques jours, pratiquée avant la huitième semaine, conduit chez le singe à une amblyopie durable. Cependant, une inversion des sutures pratiquée pendant cette période permet de restaurer la fonction de l’œil amblyope, ce qui constitue une démonstration de la plasticité cérébrale. Les observations cliniques permettent d’extrapoler ces données pour proposer que la période sensible du système visuel à une perturbation de la qualité de l’image s’étend chez l’homme jusque vers 5 ans avec un pic de sensibilité entre 6 et 18 mois. Il faut cependant distinguer la période sensible des différents éléments de la fonction visuelle : la fonction binoculaire est plus précoce que la fonction d’acuité. Certaines fonctions comme la vision des couleurs n’ont pas de période sensible.
II – Le décodage du sens : le niveau gnosique
C'est le cortex qui assure les traitements de haut niveau, débouchant sur l'interprétation de ce qui est vu, et l'accès à la signification.
Selon le type d'information, le décodage s'effectue dans des zones anatomiquement distinctes des lobes occipitaux. On peut donc distinguer cliniquement des "modules séparés, indépendants" qui traitent (fig 4) :
- Chez l'enfant plus grand, les pictogrammes, l'écriture.
 |
| Figure 3 : Les voies visuelles : organisation fonctionnelle. (Modifiée d'après MAZEAU 1995) |
CONCLUSION
Les voies neurovisuelles, constituent un ensemble fonctionnel illustrant les traitements d'informations spécifiques par "module" au niveau cérébral.
Chez l'enfant, le fait que la maturation des différentes voies visuelles n'est pas homogène et s'effectue avec des rythmes différenciés, complique singulièrement l'étude de ces modules et par là même, l'examen clinique d'un nourrisson atteint d'une déficience visuelle ou d'une forte amétropie. La figure 4 résume l'organisation fonctionnelle des voies visuelles. Une pathologie visuelle peut agir sur tout ou partie de ces voies.
CHAPITRE 3 : ACUITE VISUELLE CHEZ UN ENFANT D’AGE PREVERBAL
Introduction
Les deux méthodes de mesure de l'acuité visuelle les plus utilisées chez le très jeune enfant
· des méthodes comportementales telle la méthode du regard préférentiel,
· des méthodes objectives : poursuite visuelle de mires structurées, PEV, plus rarement le nystagmus optocinétique.
I- Méthodes comportementales subjectives de mesure du pouvoir séparateur
1.Technique du regard préférentiel ou « Bébé vision »
- En 1958 FANTZ en utilisant des motifs comme des triangles, une croix et un cercle, un damier, a pu montrer une évolution de la vision du nourrisson avec l'âge. Il a mis en évidence une caractéristique du système visuel, présente dés la naissance, qui consiste en un intérêt préférentiel et automatique pour les stimuli comportant un maximum de contrastes et de contours, alors que des stimuli lisses homogènes et uniformes provoquent au contraire un désintérêt de l'enfant.
- En 1962 ce même auteur améliore ses motifs et présente des rayures noires et blanches. Cette technique permet la mesure du pouvoir séparateur de l'oeil à l'aide de tests de différentes fréquences spatiales, imagés par des réseaux. Ce test ne sollicite pas de réponse verbale, par opposition à l'identification d'un dessin tel que dans l'échelle de SANDER-ZANLONGHI (ZANLONGHI 1999) qui fait appel à la
Cette technique dite du regard préférentiel offre pour intérêt principal de comparer les valeurs
trouvées à des valeurs dites normales pour l'âge d'une part, et d'autre part de mettre en évidence une différence d'acuité inter-oculaire.
a) Cartons de Teller
Ce test utilise le principe du regard préférentiel [TELLER 1974]. Au départ la méthode de mesure est à choix forcé; l'examinateur ignore l'emplacement du stimulus structuré et ne peut pas voir le reflet cornéen. En 1985 apparaît une méthode simplifiée avec l'utilisation de cartes d'acuité qui ont l'avantage de jouer avec l'enfant à travers un " théatre " pour attirer son regard, de réaliser en quelques minutes un examen fiable, d'être un examen simple peu encombrant et peu onéreux .
Les tests sont constitués de cartons rectangulaires gris uniformes, comportant un carré latéralisé de 12,5 cm de côté, muni de rayures noires et blanches calibrées, numérotés de 15 à 1 et présentés à une distance de 38 centimètres à 3 mois, 55 centimètres à 6 mois et 80 centimètres à 12 mois. Un sous-ensemble d'une raie noire
et d'une raie blanche s'appelle un cycle. Chaque carton est caractérisé par le nombre de cycles par centimètre de son réseau. La progression d'une planche à l'autre se fait par demi-octave, l'octave étant le double ou la moitié de la fréquence spatiale, c'est-à-dire du nombre de cycles par centimètre du réseau.
En tenant compte de la distance de présentation des tests, on convertit les cycles par centimètre en cycles par degré, ce qui représente la valeur en degrés de l'angle visuel qui sous-tend d'une part l'image rétinienne et, d'autre part, l'objet regardé et on calcule ainsi l'acuité visuelle correspondante. Ainsi, par exemple pour une présentation des cartes à 57 cm : la carte 5 de 9,8 cycles par centimètre est égale à 20/15 de vision, ce qui est supérieur à 3/10ème d'acuité visuelle (tableau 1).
 |
| Tableau 1 Tableau de correspondance entre l'acuité visuelle notée en décimal et la fréquencespatiale des cartes pour 3 distances d'examen (cartes de TELLER ) |
On peut considérer comme normale l'acuité visuelle aux cartons de Teller, si elle est de 3 cycles par degré (environ 1/10ème) à 3 mois, 9 à 12 cycles par degré (environ 3-4/10ème) à 12 mois et 30 cycles par degré soit environ 10/10ème à 4 ans (figure 5 et tableau 2).
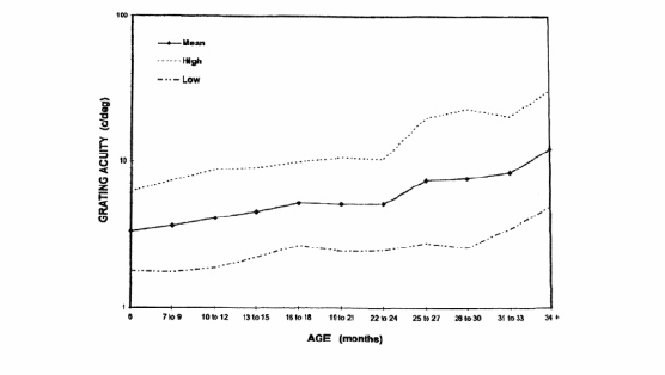 |
| Figure 5 Linéarité du développement de l'acuité visuelle binoculaire chez 1059 enfants normaux âgés de 6 à 34 mois : mesures réalisées avec les cartes de Teller [SPIERER 1999]. |
 |
|
Tableau 2 : Normes en fonction de l'âge pour les cartes de TELLER en binoculaire et en monoculaire [SALOMAO 1995] |
b) Le bébé vision tropique (BVT)
Ce test a été crée en France par François Vital-Durand [VITAL-DURAND 1996]). Les cartes d'acuité du bébé vision tropique sont des panneaux légers de 55 sur 25 cm au fond gris uniforme. Une des moitiés porte un motif circulaire concentrique dont la densité suit un profil sinusoïdal atténué sur les bords. La luminance moyenne du motif est identique à celle du fond et constante dans toutes les cartes. Le contraste maximum du motif est élevé, proche de 100 %.
La série comporte treize cartes dont les fréquences spatiales couvrent une gamme de 0,28 à 14 cycles par centimètre (tableau III). L'intervalle choisi entre les cartes successives est basé sur une échelle logarithmique. Il est égal à 1/3 d'octave pour la partie de la gamme la plus utilisée en clinique : de 2,8 à 14 cycles par centimètre. Pour les fréquences inférieures, l'intervalle est de 2/3 d'octave (Fig 6).
 |
|
Figure 6 Principe de regard préférentiel. Exemple de 5 cartes de BVTropique (Collection VITAL-DURAND INSERM Lyon) |
 |
|
Tableau 3 : tableau de correspondance entre l'acuité visuelle notée en décimale et la fréquence spatiale des cartes pour 3 distances d'examen (cartes BVT ) |
c) Résultats de la technique du regard préférentiel
Avec cette technique, la résolution spatiale s'améliore de la naissance où elle est proche de 1 cycle par degré (20/600 soit 0.3/10), à 15 cycles par degré (20/40 soit 5/10) vers la fin de la première année [ATKINSON 1982, DOBSON 1990 , TELLER 1979, TELLER 1986, VITAL-DURAND 1992].
Une étude sur 500 examens d'acuité visuelle du nourrisson avec les cartons de Teller réalisés par Vital-Durand [1990] a montré que l'on pouvait considérer chez les enfants dits normaux, une résolution de 6,5 cycles par degré soit environ 2,5/10ème à 4 mois, 9 cycles par degré soit environ 3/10ème à 8 mois et jusqu'à 12 cycles par degré soit environ 4/10ème à 1 an, en vision binoculaire. En vision monoculaire, les chiffres sont inférieurs d'une demie-octave, c'est à dire que l'on obtient respectivement 4,5 cycles par degré (1,5/10ème), 7 cycles par degré (2,2/10ème) et 9 cycles par degré (environ 3/10ème). Les valeurs obtenues chez les enfants porteurs de pathologie, sont nettement inférieures et beaucoup plus dispersées. Au cours de la première année les acuités visuelles sont sensiblement plus élevées chez les enfants nés prématurément que chez les enfants nés à terme en se référant à l'âge de conception [VAN HOF-VAN DUIN 1986].
A un âge donné la différence interoculaire est au maximum de 0,5 octave. Une différence plus importante est considérée comme un bon indicateur de baisse d'acuité visuelle monoculaire.
Par contre ce test du comportement visuel n'est pas à considérer comme une bonne méthode de dépistage de l'amblyopie unilatérale en raison du nombre important de faux positif (ésotropies précoces) ou de faux négatifs [BOURRON-MADIGNIER 1997].
2 . La poursuite visuelle d'une mire structurée en mouvement
a) Principe
Tout comme le NOC et le regard préférentiel, la poursuite visuelle d'une mire en mouvement est une méthode utilisant la réponse oculomotrice à une stimulation [BEASLEY 1933]. La mire se déplace horizontalement à partir d'une position centrale, d'une extrémité à l'autre de l'écran, selon un mouvement de va et vient (fig. 8). La poursuite fait appel à la fixation centrale. Cette technique est donc différente de celle du nystagmus opto-cinétique dans laquelle la mire se déplace toujours dans le même sens.
L'enregistrement des mouvements oculaires peut se faire soit par l'intermédiaire de l'électro-oculographie cinétique, soit par un système de caméra-vidéo, soit par la technique de détection de champ magnétique, soit par une technique de photo-oculographie différentielle ; POG (fig. 8) [BUQUET 1996]. Ces sont ces deux dernières méthodes qui sont actuellement utilisées en clinique : la POG chez le nouveau né et l'électro-oculographie cinétique chez l'enfant plus grand.
 |
|
Figure 7 Dispositif de poursuite de mire par photo-oculographie (bébé âgé de 7 jours) (Collection M. CHARLIER, INSERM Lille) |
b) Résultat
La mesure du pouvoir séparateur est donnée par la taille du plus petit test que l'enfant parvient à suivre du regard pendant au moins 2 allers et retours [CHARLIER 1993]. Les mouvements des yeux sont enregistrés parallèlement au déplacement de la mire .
 |
Figure 8 : tracé chez un nouveau né sans pathologie de 4 jours obtenu par photo-oculographie. Les tracés obtenus sont des tracés de poursuite oculaire avec 2 voies, l'une correspondant à la composante horizontale du mouvement, l'autre à la composante verticale. (Collection J. CHARLIER, INSERM Lille) La poursuite visuelle de mires structurées a fait la preuve de son intérêt sur une série de plus de 500 enfants [DEFOORT-DHELLEMMES 1984 , TOURNIER 1991] et chez le nouveau né. Son interprétation est néanmoins moins facile chez le nouveau né dont la poursuite est entrecoupée de saccades de correction [BUQUET 1992]. La vitesse de déplacement de la mire doit être lente et ne pas dépasser 19 degrés/seconde chez les nouveau-nés [KREMINITZER 1979]. Ces saccades ont tendance à diminuer avec l'âge. |
II – Méthodes comportementalesobjectives du pouvoir séparateur
Le nystagmus optocinétique (NOC)
a) Définition :
Le NOC se caractérise par une série de mouvements oculaires réflexes se produisant de façon répétée chez un sujet qui regarde une succession d'objets défilant rapidement devant ses yeux (figure 9).
 |
Figure 9 Principe du NOC. Un enfant de 7 mois est placé entre 30 et 80 cm du tambour sur lequel figure des mires soit sous formes de barres noires et blanches verticales soit, comme ici, sous la forme de figure et de dessin. (Collection M. VITAL-DURAND, INSERM Lyon) Lors du NOC, deux types de mouvement vont se succéder alternativement tant que dure la stimulation : un premier mouvement de poursuite lent et uniforme dans le sens du déplacement des objets, un deuxième mouvement à type de saccades qui ramènent brutalement les yeux à leur position de départ (figure 10). |
 |
|
Figure 10: Exemple de tracé de NOC chez un bébé avec une vision normale âgé de 3 mois (Collection M. CHARLIER, INSERM Lille) |
b) Principe :
Une succession régulière de tests (soit des bandes alternativement noire et blanche, soit des dessins) défile devant les yeux de l'enfant dans une direction et à une vitesse donnée. Le pouvoir séparateur équivaut à la valeur angulaire du plus petit test qui va introduire un NOC.
c) Résultat :
Ce test est le plus couramment utilisé pour donner une information globale sur la présence ou l'absence de la fonction visuelle chez un patient non coopérant ou un enfant. On peut considérer que si l'on obtient une réponse positive avec une certaine largeur de bande, le sujet est capable de détecter un stimulus qui lui a été présenté. Par contre, si le nystagmus optocinétique n'est pas interprétable chez un nouveau né, on ne peut établir le fait que l'enfant ne voit pas . Historiquement c'est la première méthode utilisée chez le bébé pour mesurer le pouvoir séparateur : Gormann [1957 ] donne la valeur de 1,0 cpd pour 80 % des bébés de moins de 5 jours et Dayton [1964 ] donne 1,5 cpd pour 50 % des bébés âgés de moins de 8 jours.
En théorie, on peut considérer que des bandes très fines donnant lieu à un nystagmus optocinétique indiquent la limite de la résolution spatiale de l'enfant. En établissant une relation entre la largeur de la bande et l'angle visuel qu'elle sous-tend, certains chercheurs ont cherché à évaluer l'acuité visuelle [ENOCH 1976, FANTZ 1962] ; celle-ci, qui fait intervenir le mouvement d'un test, est différente de l'acuité visuelle angulaire obtenue avec le test
traditionnel statique des E de Snellen. Les résultats sont résumés dans le tableau 4

Tableau 4 : valeur d'acuité visuelle obtenue par NOC à différents âges.
En conclusion, une réponse à l'enregistrement optocinétique signifie que la fonction visuelle est présente. En corollaire, cette méthode de mesure donne une bonne information qualitative de la fonction visuelle du bébé, mais elle est plus difficile à utiliser pour obtenir une acuité visuelle " chiffrée ".
III – Acuite visuelle et maturation par potentiels évoqués visuels
L'acuité visuelle peut être évaluée en fonction de l'amplitude du PEV damiers et de la fréquence spatiale du stimulus visuel utilisé (fig. 11). Chez l'adulte, une courbe de référence a été établie à partir de l'étude de sujets normaux [HACHE 1979 ]. Cette courbe d'amplitude en fonction de la fréquence spatiale fait un large U inversé. Elle est maximale pour des fréquences spatiales proches de 2 cycles par degré (15 à 20').
 |
|
Figure 11 bébé de 4 mois placé devant un écran de stimulation avec des damiers de 60' présentés à une distance oeil-écran de 1 mètre. (Collection Dr DHELLEMMES-DEFOORT Lille) |
Idéalement, l'acuité visuelle pourrait être mesurée en utilisant des tests de fréquence spatiale croissante, et l'acuité visuelle estimée comme la valeur de la fréquence spatiale du plus petit test qui donne une réponse. En pratique ce n'est pas réalisable car plus on se rapproche du seuil de perception qui correspond à la meilleure acuité visuelle, plus l'amplitude du PEV est faible.
1. Par extrapolation [REGAN 1977 ]
Pour évaluer le pouvoir séparateur d'un enfant, il est nécessaire d'utiliser des tests de différentes fréquences spatiales (au moins cinq chez l'enfant de plus de 1 an) et de construire la courbe amplitude de la réponse évoquée visuelle en fonction de la fréquence spatiale.
Une droite de régression est construite à partir des valeurs d'amplitude des pics de haute fréquence spatiale enregistrables (3 fois supérieurs au bruit de fond). L'intersection entre cette droite de régression et l'abscisse (amplitude zéro) donne une valeur de fréquence spatiale qui correspond au pouvoir séparateur. (Fig.12)
Cette méthode de mesure de l'acuité peut être utilisée avec un mode de stimulation transitoire.
Le mode stable plus rapide, lui est préféré en clinique, surtout dans les pays anglo-saxons.
Chez l'enfant, la technique employée est le "PEV par balayage " ou (sweep PEV).
Avec cette méthode, les mesures d'acuité chez l'enfant normal sont reproductibles à 0.5 octave près (une octave correspond à une variation de fréquence spatiale d'un facteur 2). La différence d'acuité entre les 2 yeux est inférieure à une octave.
Figure 12 Exemple de " sweep PEV ". L'intersection entre la droite de régression et l'abscisse(amplitude zéro) donne une valeur de fréquence spatiale qui corres pond au pouvoir séparateur . (modifié d'après [Norcia 1985])

2. Les facteurs de variation de l'acuité visuelle par PEV chez l'enfant
a) Age
L'âge joue un rôle primordial. Il est démontré que le pouvoir séparateur mesuré par PEV progresse rapidement de la naissance jusqu'à l'âge de 6-8 mois que ce soit avec des stimulations transitoires ou stables, que ce soit avec des barres noires et blanches alternantes, ou avec un profil sinusoïdal [Harris 1976 , Norcia 1985 , Norcia 1987 , Tyler 1979 ], ou des damiers alternants [SOKOL 1978 ], ou bien une apparition-disparition de damier [De Vries-
Les valeurs d'acuité visuelle sont résumées dans le tableau 6.
b) Influence de la fréquence temporelle
L'amplitude du PEV varie avec la fréquence temporelle du test (qui est égale à la moitié du nombre de renversements par seconde).La fréquence temporelle maximale à laquelle Fiorentini [1992 ] enregistre un PEV avec un test de 60' est 6 Hz entre 1 et 2 mois, 8 Hz entre 2 mois et 3 mois et demi.
c) L'accommodation et la convergence
La focalisation de l'image rétinienne n'intervient pas en PEV par damiers chez les enfants de moins de 2 mois, qui ont un pouvoir de résolution spatiale bas [Atkinson 1977, SALAPATEK 1976 , Sokol 1983 ].
d) Influence de la réfraction
Les anomalies de réfraction doivent être parfaitement corrigées (myopie et astigmatisme en particulier) chez l'enfant comme chez l'adulte.
IV – Comparaison des différentes méthodes de mesure du pouvoir séparateur chez le jeune enfant
Les tableaux suivants (5 et 6) donnent une estimation du pouvoir séparateur obtenu par PEV, par la technique du regard préférentiel, par la poursuite, puisque nous avons vu que le NOC ne donne qu'une valeur qualitative.
 |
| Tableau 5 : Acuité visuelle obtenue chez les nouveau-nés âgés de moins de 9 jours. |
 |
|
Tableau 6 : estimation du pouvoir séparateur obtenu par PEV et par regard préférentiel chez le jeune enfant de 1 mois à 4 ans (* acuité monoculaire moyenne). |
Les valeurs d'acuité obtenues par les PEV sont meilleures que celles obtenues par la méthode subjective : le regard préférentiel.
Cette différence fait intervenir plusieurs hypothèses :
* Les PEV par petits damiers (15') correspondent à l'activité de l'aire fovéolaire (2°),
alors que le regard préférentiel étudie la rétine plus périphérique.
* Chaque technique étudie un stade différent du processus neuronal.
* Les PEV étudient la transmission du message visuel au cortex visuel, alors que le regard préférentiel fait intervenir l'intégration du message visuel au niveau des aires associatives.
* La méthode du regard préférentiel fait appel à la motricité de la tête et des yeux. Elle a le désavantage d'être influencée par l'immaturité des aires motrices.
* Les critères d'évaluation de l'acuité visuelle sont différents. Ils sont considérés comme plus sévères dans la méthode de regard préférentiel où l'acuité est estimée comme la taille du plus petit test qui donne 75 % de bonnes réponses par l'observateur. En utilisant des critères plus généreux (vers 55-60 % de bonnes réponses) l'acuité visuelle par la méthode de regard préférentiel s'améliore de 1 à 1,5 octaves et approche les valeurs obtenues par les PEV [Dobson 1978 ].
* Les propriétés temporelles des stimulus sont différentes. Avec les PEV, il existe une variation temporelle du stimulus, pas avec le regard préférentiel. L'acuité visuelle mesurée par la méthode de regard préférentiel est meilleure si on utilise des tests alternants à une fréquence de 14 renversements par secondes au lieu des tests statiques [SOKOL 1982 ].
Toutefois, l'acuité visuelle obtenue avec des PEV de même fréquence temporelle reste meilleure de 2 octaves à 2 mois, de 0,5 octave à 12 mois.
CONLUSION
- La méthode de regard préférentiel est utile pour suivre la maturation et l'amélioration de l'acuité visuelle du nourrisson. C'est un complément utile mais insuffisant pour le dépistage de l'amblyopie unilatérale [SPEEG-SCHATZ 1995 ].
- La méthode de poursuite de mires structurées est une méthode plus rapide et plus objective, mais elle permet seulement le dépistage des amblyopies importantes. Cette technique est appelée à évoluer avec des écrans de haute résolution.
- L'évaluation de l'acuité visuelle par les PEV ne peut pas constituer un test de dépistage de l'amblyopie ou de suivi régulier de l'amélioration de l'acuité visuelle après un traitement d'amblyopie. Par contre, les PEV sont indispensables chez tous les enfants ayant un comportement visuel anormal, isolé ou associé à une pathologie neurologique.