
L'adaptation thermophile

On a choisi de présenter ici les caractéristiques adaptatives suivant trois parties qui suivent l'analyse classique de l'extérieur vers l'intérieur de la cellule, de la membrane cellulaire - ou la paroi quand celle-ci est présente - frontière entre la cellule et son environnement, vers le coeur du cytoplasme et ce qui constitue la mémoire évolutive de la cellule, son information génétique. On considère ici uniquement des organismes unicellulaires procaryotes (Bactéries et Archées).
- l'adaptation thermophile au niveau de la membrane1,
- l'adaptation thermophile au niveau du cytosol (principalement les activités enzymatiques et la structure des protéines)2,
- l'adaptation thermophile au niveau génétique3.
Toutefois il est essentiel de retenir que cette adaptation thermophile est un tout et met en oeuvre simultanément les différentes stratégies exposées. Ce qui n'est pas sans poser des difficultés pour comprendre comment une telle adaptation globale, complexe, multiple, a pu se mettre en place au cours de l'évolution du monde vivant.
"les bioextrémistes du thermomètre" : où l'on trouve aux deux extrêmes du "thermomètre du vivant" des Archées ; mais elles ne sont plus en réalité tout à fait aussi seules qu'on l'avait initialement supposé ou qu'on continue à le laisser croire... Toutefois si des Bactéries les ont rejointes voire dépassées dans le domaine du grand froid (les psychrophiles et cryophiles), elles restent (au moins jusqu'ici) les championnes pour le domaine hyperthermophile dont elles détiennent le record.
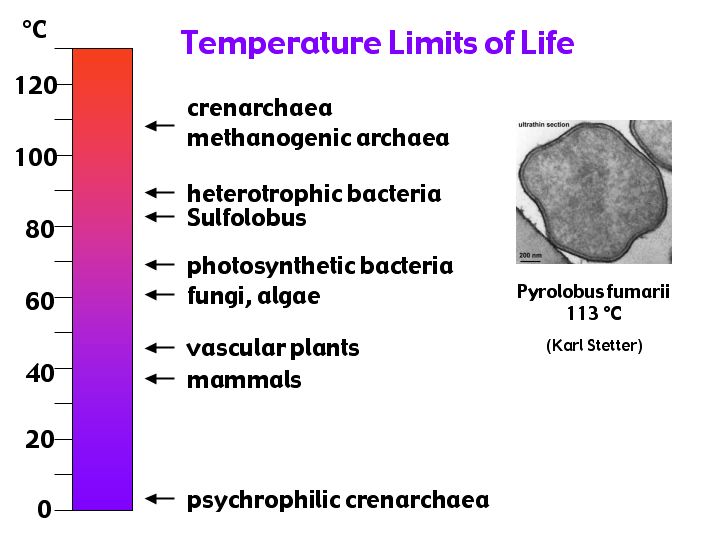{kind=link}
L'adaptation membranaire (voire pariétale)
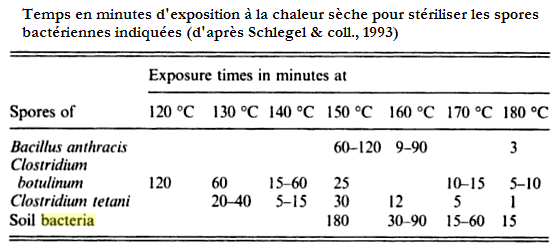
Une première idée pouvant venir à l'esprit (naïf!) serait que les microorganismes thermophiles possèdent une paroi constituant le bouclier miracle qui les aurait protégés de la terrible chaleur. En fait non, les thermophiles et hyperthermophiles ne sont pas particulièrement mieux dotés de paroi que leurs apparentés non thermophiles. Toutefois on ne peut oublier par ailleurs le rôle joué par de telles parois dans, par exemple , la résistance à la chaleur des spores de nombreuses bactéries. On doit aussi souligner que dans le cas particulier des organismes thermoacidophiles qui cumulent en quelque sorte les inconvénients d'un milieu très chaud et très acide (inconvénient dont ils font un avantage!), les Archées [à vérifier qu'il n'y a pas aussi des bactéries] que l'on y rencontre sont très généralement pourvues d'une paroi particulièrement épaisse. Généralement, car il y a aussi des organismes comme Thermoplasma qui s'en sortent sortent tout aussi bien dans le même milieu, et sans paroi!
{kind=link}
C'est donc bien pour l'essentiel au niveau de la membrane qu'est gérée cette difficulté. Et la solution trouvée tient pour l'essentiel dans les caractéristiques des lipides membranaires. On sait encore peu de choses sur les protéines membranaires, nous n'en parlerons pas (ou plus exactement on retiendra que les adaptations de ces protéines assurant les échanges avec l'environnement doivent être comparables à celles que nous décrirons plus loin pour les enzymes thermophiles, encore que la membrane soit un compartiment un peu différent du cytoplasme...). Il n'a pas à notre connaissance été décrit d'autres mécanismes d'adaptation au niveau membranaire que ceux impliquant les lipides (leur longueur, la nature des liaisons chimiques, leur degré d'insaturation, leur conformation, les modifications par des radicaux, etc. : tableau ci-après).
Toutes les adaptations présentées peuvent se résumer à l'idée suivante : il faut rigidifier la membrane pour empêcher les fuites (eau, ions) que tend à provoquer naturellement l'élévation de la température. Et de fait à température plus douce, à température moyenne, ces membranes ne sont plus fonctionnelles car trop rigides. On peut d'ailleurs ajouter que c'est exactement l'inverse qui se produit pour les microorganismes psychrophiles adaptés au froid : tout est fait pour fluidifier la membrane alors que l'abaissement de température tend à la rigidifier.
| adaptation au froid | adaptation à la chaleur |
| promouvoir la fluidité de la membrane alors que la température est basse | réduire la fluidité de la membrane alors que la température est très élevée |
| insaturation des lipides (nombreuses doubles liaisons) | saturation des lipides (rares doubles liaisons) |
| doubles liaisons en position cis | doubles liaisons en position trans |
| chaînes plus courtes | chaînes plus longues |
| chaînes ramifiées (methyl-branching) | chaînes droites (straight chain) |
Une illustration grossière mais utile de ces principes consiste à comparer les comportements au congélateur, au réfrigérateur et à température ambiante voire en chauffant, d'une huile végétale, de beurre, de margarine ou encore d'une graisse animale. La mise en relation avec la composition en acides gras de ces différents produits explique les différences de comportement et montre l'importance des propriétés énumérées dans le tableau ci-dessus. C'est en effet un fait d'alimentation bien connu que les matières grasses végétales sont plus riches en lipides insaturés et même polyinsaturés. C'est cette différence de composition qui explique l'abaissement du point de fusion (ou de solidification - selon que l'on regarde la transformation dans un sens ou dans l'autre).
Des lipides de longueur inhabituelle
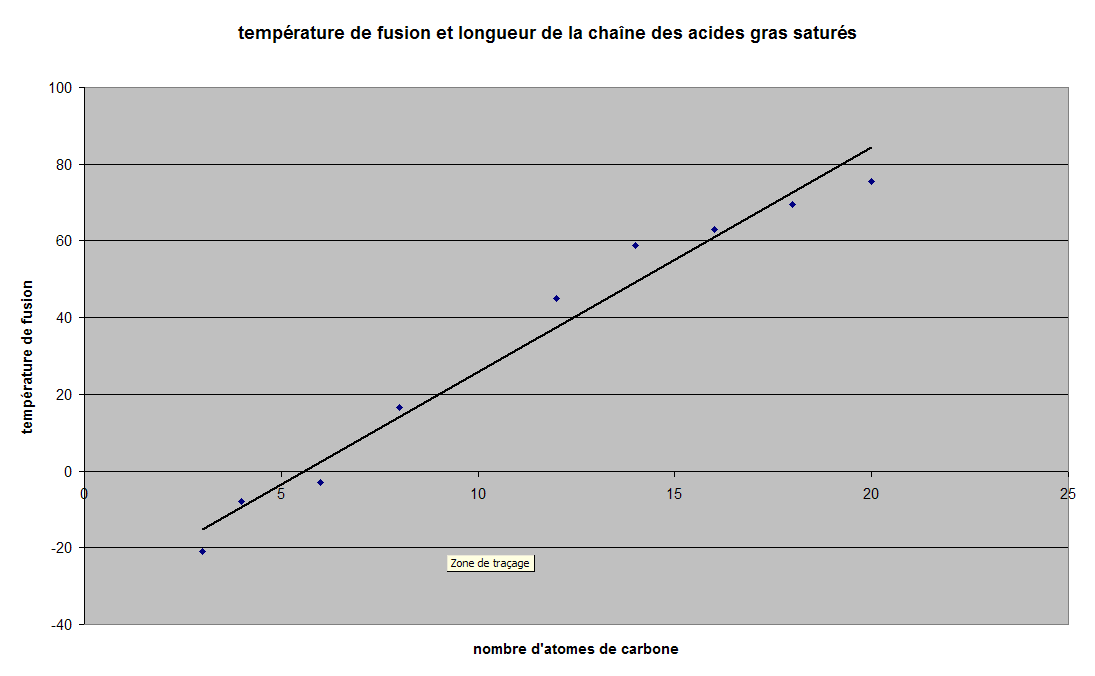
Un constat effectué initialement chez les Archées est la présence de lipides d'une longueur exceptionnelle qui traversent toute l'épaisseur de la membrane et contribuent ainsi à former une monocouche et non la bicouche classique qui a l'inconvénient de se séparer en son milieu lors de l'exposition à haute température. Ces lipides qui traversent toute la membrane contribuent à sa rigidification. Il existe une convergence remarquable pour cette propriété entre Archées thermophiles et Bactéries thermophiles. Des transferts génétiques horizontaux - des transferts des gènes de biosynthèse de ces lipides - entre espèces non apparentées, seraient en cause.
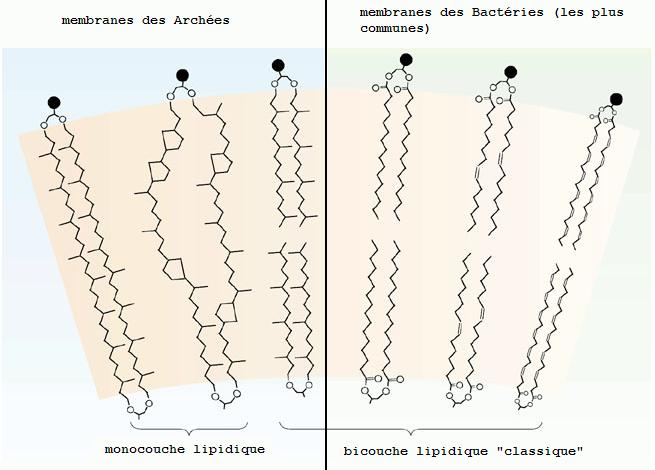{kind=link}
|
Des lipides membranaires originaux chez les Archées thermophiles |
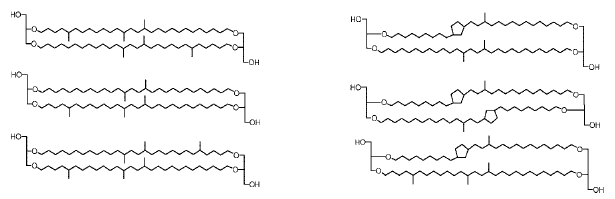 |
|
et leurs équivalents chez les Bactéries thermophiles (d'après Schouten & coll.) |
|
On peut ici pour compléter cette caractéristique rappeler que plus la longueur d'un acide gras augmente et plus sa température de fusion sera faible. Ainsi pour une température donnée, une réduction de longueur des acides gras tend à accroître la fusion et donc l'état liquide, plus fluide. A l'inverse, un allongement de acides gras tend à abaisser leur point de fusion, donc à les déplacer vers l'état solide, plus rigide.
{kind=link}
Des liaisons chimiques inhabituelles
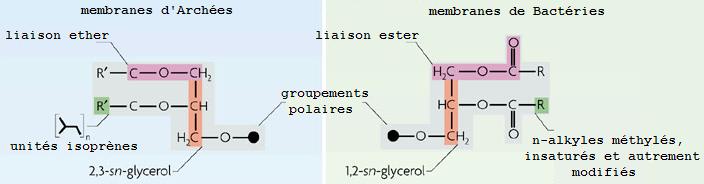
La liaison covalente qui relie les chaînes d'acides gras au glycérol est de type éther chez les Archées alors qu'elle est de type ester classiquement, ce que l'on retrouve par exemple chez les Bactéries. La liaison ether est plus stable et résiste mieux aux températures élevées.
{kind=link}
Conformations cis/trans et degré d'insaturation
|
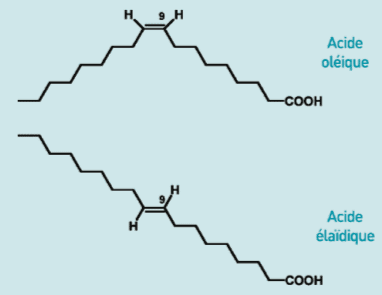
Une double liaison en position cis déforme considérablement la chaîne carbonée, réduit le point de fusion et tend donc à accroître la fluidité de la membrane. Au contraire une double liaison trans préserve une structure linéaire, réduit dans une moindre mesure la température de fusion et contribue davantage que la précédente à une relative rigidité. |
 |
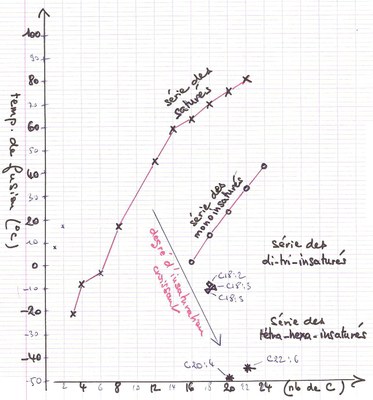
On l'a rappelé plus haut, les acides gras insaturés favorisent la fluidité de la membrane alors que les saturés en augmentent la rigidité. C'est ce dernier point qui est mis à profit par les Thermophiles. Le graphique ci-dessous illustre la relation entre température de fusion et la longueur et le degré d'insaturation de la chaîne carbonée des acides gras (d'après des sources diverses dont wikipedia).
Des modifications par addition de substituants
C'est chez les organismes thermophiles l'absence de modifications - et donc de ramifications et "d'encombrements latéraux" autour de la chaîne d'acide gras - qui est favorable à la rigidité du lipide et ainsi de la membrane dans laquelle il s'insère. On peut citer en particulier l'absence de méthylations, une modification secondaire que l'on pourra au contraire rencontrer chez les psychrophiles adaptés au froid.
L'adaptation au niveau de l'organisation et du fonctionnement du cytosol/cytoplasme
La composition chimique du cytosol
On songe ici à des adaptations comme on en connaît dans le domaine de l'adaptation au froid avec les substances de type "antigel". Il existe bien des composés (ions minéraux, petites molécules biologiques comme la bétaïne) intervenant dans la régulation osmotique chez les halophiles des milieux hypersalins. Mais dans le domaine de la thermophilie il n'y a eu semble-t-il jusque là aucune description montrant clairement l'implication de composés thermoprotecteurs spécifiques ayant de telles propriétés. A une exception près toutefois puisqu'une piste de travail a été ouverte concernant un dérivé de l'inositol.
L'activité enzymatique et la structure des protéines
Au contraire dans le domaine de l'enzymologie (et des protéines en général) de nombreux travaux ont très clairement identifié des stratégies d'adaptation aux hautes températures. Les principes généraux dégagés des comparaisons entre protéines spécifiques ou protéomes complets d'organismes mésophiles et thermophiles apparentés sont les suivants :
- des surfaces externes (au contact du solvant) fortement chargées ; les protéines d'organismes thermophiles présentent en surface davantage de résidus chargés d'acides aspartique et glutamique, d'arginine et de lysine ;
- une conformation plus rigide maintenue par un réseau de multiples interactions ioniques, avec les mêmes acides aminés que ci-dessus mais cette fois interagissant deux à deux (un acide aminé acide - acide aspartique ou acide glutamique - lié à un acide aminé basique - lysine ou arginine) sous la forme de paires d'ions à énergie de stabilisation forte ;
- un coeur hydrophobe fortement compacté (dont des acides aminés hydropobes à chaîne latérale en moyenne plus courte),
- une tendance générale vers une structure plus compacte obtenue en minimisant les vides, en éliminant les boucles par exemple.
Pour autant la structure tridimensionnelle des protéines reste très généralement la même que les organismes soient psychrophiles, mésophiles ou thermophiles. Bien entendu parce que c'est cette structure qui donne sa fonction à la protéine et que les gènes concernés sont essentiellement des homologues traduisant la parenté commune de tous ces organismes.
Il semble aussi que - dans de rares cas - il ait pu y avoir au cours de l'évolution substitution d'une protéine par une autre (substitution de gènes plus exactement) qui aura par convergence acquis la même fonctionnalité.
L'expression de protéines recombinantes thermostables chez des hôtes mésophiles a, pour sa part, très clairement montré que la protéine portait en elle l'essentiel des déterminants de sa thermostabilité.
Cependant on observe dans un certain nombre de cas des anomalies de conformation qui suggèrent que des voies particulières d'établissement d'une conformation correcte des protéines thermophiles existent spécifiquement chez les organismes thermophiles. Ces voies impliquent les protéines de choc thermique (inductibles) et les protéines chaperon(e)s. Toutefois certaines parmi celles-ci sont néanmoins absentes chez des organismes hyperthermophiles! A l'évidence une situation difficile à analyser que les recherches en cours devraient permettre d'éclaircir.
La thermostabilité des acides aminés
C'est là le point majeur permettant de comprendre la thermostabilité des protéines. Les comparaisons entre couples de protéines ou couples d'ensembles de protéines (qualifiés de protéomes) thermophiles vs. mésophiles a clairement montré comment certains acides aminés étaient "évités" (ils sont moins fréquents dans les protéines thermophiles) alors que d'autres étaient "recherchés" (ceux-là sont au contraire plus fréquents). Un certain nombre d'indices de thermostablité ont ainsi été proposés par différents auteurs. Nous proposons dans ce dossier une activité sur ce thème.
Les acides aminés qui apparaissent dans ces comparaisons les plus contre-sélectionnés sont l'asparagine et la glutamine d'une part, la sérine et la thréonine d'autre part, dont les fonctions amide et hydroxyle respectivement sont instables à haute température. Les acides aminés qui sont au contraire fortement sélectionnés, "favorisés", sont l'acide glutamique, l'arginine, la lysine, et la tyrosine dans une moindre mesure qui ont tous l'avantage d'être des acides aminés longs permettant des interactions de type hydrophobe tout le long de la chaîne latérale et qui sont en outre des acides aminés chargés pour les trois premiers. Notons que l'abondance d'acides aminés basiques tend à donner en moyenne des points isoélectriques basiques aux protéines thermophiles.
|
L'indice proposé ici a été établi à partir de plusieurs indices trouvés dans la littérature scientifique :
|
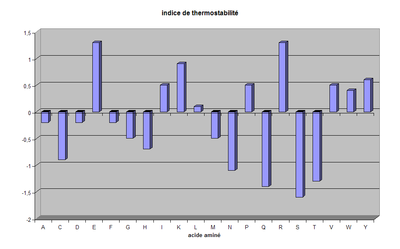 |
La polymérisation des sous-unités
Il est fréquemment fait mention du fait que chez les thermophiles on observe plus souvent des enzymes à l'état oligomérique là où les équivalents mésophiles sont à l'état de monomère, ou respectivement des formes tétramériques chez les thermophiles là où s'observent des formes dimériques chez les mésophiles. Toutefois il n'y pas eu à notre connaissance de travail de synthèse conséquent analysant de façon critique ce sujet.
La compartimentation cellulaire
Elle constitue manifestement un obstacle important à l'adaptation thermophile et aucun organisme à compartimentation cellulaire - qu'il s'agisse d'eucaryotes (tous les Eucaryotes) ou de procaryotes (par exemple les Cyanobactéries) - n'entre dans la catégorie des hyperthermophiles. Toutefois quelques Eucaryotes et quelques Cyanobactéries trouvent leur place au sein des thermophiles, ou tout au moins des sub- ou quasi-thermophiles selon que l'on fixe la "barre" de la thermophilie à 45, 50 ou 60°C.
L'adaptation génétique
Les particularités des chromosomes
Que ce soient par la forme, le nombre, la taille, la structure des chromosomes et de la chromatine, ou les protéines associées à l'ADN, il ne semble pas y avoir de fait marquant à mettre en relation avec le statut thermophile des organismes.
Il y a toutefois une particularité au niveau de l'ADN, qui est la présence d'un surenroulement positif (= dans le sens de la rotation, qui ferme la double hélice d'ADN et la stabilise à haute température). Ce surenroulement positif est une spécificité des organismes thermophiles (et des bactéries hyperthermophiles), tous les autres organismes présentent un surenroulement négatif de la molécule d'ADN (=dans le sens inverse de la rotation de la double hélice d'ADN, et qui favoriserait la séparation des 2 brins). Cette particularité est due à la présence d'une enzyme chez les thermophiles, une topoisomérase de type I appelée la gyrase réverse. La reverse gyrase est capable de surenrouler positivement les ADN circulaires covalemment fermés. On notera aussi que chez certaines Archées sont observées des protéines histone-like ce qui les rapproche considérablement de ce point de vue des organismes eucaryotes.
Les particularités du génome et de l'ADN
Les thermophiles - et plus encore les hyperthermophiles - se caractérisent par des génomes de plus petite taille, l'essentiel du "gain de place" (établi par comparaison avec des espèces apparentées non thermophiles) se faisant en réduisant la taille des espaces intergéniques. En effet, le nombre de gènes n'est pas fondamentalement très différent et la taille des gènes tout au plus légèrement réduite parfois. En outre, une réduction dans les familles de gènes (les paralogues) est également régulièrement constatée, un seul gène pouvant être présent là où les espèces mésophiles proches pourront en posséder plusieurs. On le voit en particulier pour les gènes ribosomiques, les gènes des ARNt. Certains gènes pouvant parfois être totalement absents (on l'a déjà mentionné ci-dessus à propos des gènes de certaines protéines de choc thermique).
Un facteur initialement mis en avant, la composition en bases avec un fort %G+C, s'est avéré par la suite ne pas être toujours vérifié. Il garde néanmoins sa signification lorsqu'on s'adresse aux ARN.
La stabilité thermique des ARN
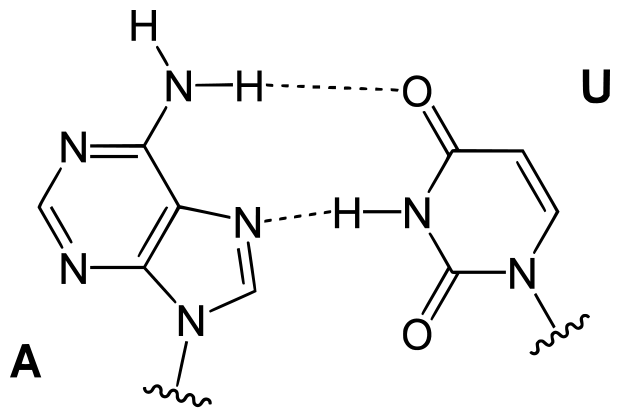
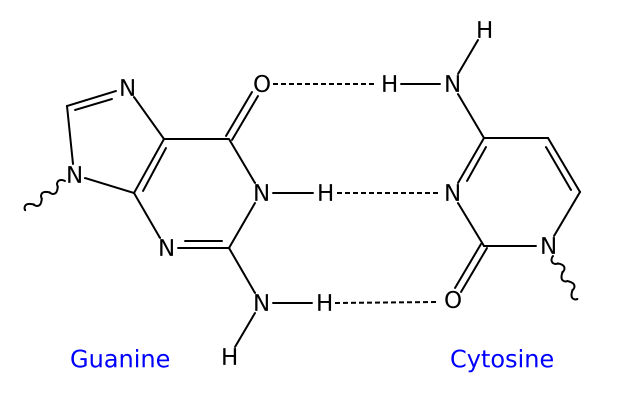
De nombreux travaux dont ceux de Galtier et Lobry (1997) [qui nous ont inspiré une activité présentée plus loin], montrent une particularité de composition en bases des ARN (qu'ils soient ribosomiques ou de transfert) : une plus grande richesse en Guanine et Cytosine (%G+C plus élevé). Cet enrichissement concerne plus particulièrement les portions double brin de ces molécules et sont indépendantes du %G+C de l'ensemble du gènome. On peut dès lors interpréter ces faits en s'appuyant sur le constat bien connu que les paires de bases G et C forment entre elles 3 liaisons hydrogène, alors que les bases Adénine et Thymine (ou Uracile pour l'ARN) appariées n'en forment que 2. Or, l'élévation de température tend à défaire ces liaisons faibles. Ainsi l'enrichissement en G+C peut conduire à rendre plus stables les structures secondaires et tertiaires des ARN, structures qui sont essentielles à leur activité.
{kind=link}
{kind=link}
|
Modèle de structure d'un ARN de transfert :
|
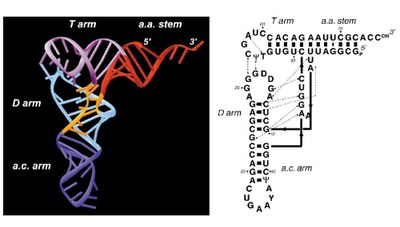 |
Une fois acquise cette relation entre %G+C des ARN (principalement la séquence de l'ARN 16S de la petite sous-unité du ribosome qui est le plus fréquemment utilisé) et température optimale de croissance de l'organisme, il est tentant de faire de cet indice ribosomique un véritable "thermomètre moléculaire" ayant une bonne valeur prédictive et pouvant être appliqué à des molécules ancestrales tirées de la comparaison des séquences de molécules actuelles. C'est ce type de démarche que les chercheurs ont engagé et il se développe ainsi une reconstitution des paléoenvironnements à l'aide des molécules biologiques actuelles. Une attention toute particulière étant portée à la reconstitution du milieu dans lequel vivait LUCA, dernier ancêtre commun à tous les organismes cellulaires actuels. Un important débat s'est développé autour de la question de savoir si LUCA était ou non un organisme thermophile voire hyperthermophile (voir par exemple les travaux du groupe de Manolo Gouy à Lyon). C'est aussi sur ce thème que nous proposons une activité [lien].
Les particularités au niveau de gènes spécifiques
Hormis ceux impliqués dans la synthèse des lipides membranaires et la gyrase réverse ou le point précédent concernant les ARN, il ne semble pas y avoir de trait particulier à souligner.
Les modalités de l'expression et la régulation génétique
Il ne semble pas là encore y avoir de trait particulier qui ait été identifié.