
Classification phylogénétique des métazoaires
Auteur : Gabriel Markov, Equipe de Zoologie Moléculaire, Institut de Génomique Fonctionnelle de Lyon, Université de Lyon, Ecole Normale Supérieure de Lyon, Université Lyon 1, CNRS, INRA, Institut Fédératif 128 Biosciences Gerland Lyon Sud, France./USM 501/UMR CNRS 5166 - Evolution des Régulations Endocriniennes. Muséum National d'Histoire Naturelle, Paris, France
Ce document constitue un aperçu de la diversité des métazoaires, non exhaustif mais suffisamment riche pour mettre en évidence l'utilité de la classification phylogénétique pour comprendre la structure de la biodiversité et aborder des questions relatives aux modalités de l'évolution. Seuls les groupes « majeurs », c'est à dire importants en termes de nombre d'espèces décrites seront présentés en détail. D'autres groupes numériquement moins importants, parfois limités à quelques espèces, seront toutefois évoqués, lorsque certains caractères observables chez eux fournissent des éléments utiles à la discussion.
Dernière mise à jour: juin 2008.
| I- Introduction | ||||
| II - La place de métazoaires parmi les êtres vivants | ||||
| III - Les "éponges", un groupe en sursis... | ||||
| IV - Les eumétazoaires | IV-A Les bilatériens | IV-A-1 Les deutérostomiens | IV-A-1-a Les ambulacraires | |
| IV-A-1-b Les chordés | ||||
| IV-A-2 Les protostomiens | IV-A-2-a Les ecdysozoaires | IV-A-2-a-1 Les arthropodes | ||
| IV-A-2-a-2 Les nématodes | ||||
| IV-A-2-b Les lophotrochozoaires | IV-A-2-b-1 Les annélides | |||
| IV-A-2-b-2 Les mollusques | ||||
| IV-A-2-b-3 Les plathelminthes | ||||
| IV-B Les cnidaires | ||||
| V- Conclusion |
I - Introduction
La classification des métazoaires (ou animaux pluricellulaires) n'est pas un simple exercice formel: elle répond à des besoins précis, qui peuvent être culinaires (classer les organismes en comestibles, non-comestibles voire vénéneux), théologiques (retrouver le plan divin qui aurait été mis en oeuvre lors de la création), ou scientifiques. Dans le domaine des sciences de la vie, il est possible de classer les animaux en fonction de critères écologiques (herbivores, charognards, fouisseurs), qui permettent de décrire le fonctionnement d'un biotope. Toutefois cette manière de classer ne tient pas compte de la dimension historique, qui suppose que les biotopes évoluent, de même que les espèces qui y vivent. La seule façon de prendre en compte la dimension temporelle, et donc de classer la biodiversité actuelle mais aussi passée, est la classification phylogénétique, qui regroupe les organismes en fonction de leur degré d'apparentement. Dans ce cadre conceptuel, on cherche à reconnaître des groupes dits monophylétiques, qui comprennent uniquement un ancêtre commun hypothétique et l'ensemble de ses descendants. La classification phylogénétique des métazoaires, encore partiellement irrésolue, constitue un ensemble cohérent d'hypothèses au sujet des relations de parenté entre les animaux et de la chronologie d'acquisition de certains traits de caractères, et ne doit donc pas être confondue avec les clés de détermination qui doivent permettre de trier de façon non ambiguë des échantillons de terrain lors d'un inventaire de biodiversité.
Longtemps demeurée irrésolue pour des raisons méthodologiques, notamment la difficulté de comparer des structures anatomiques très différentes, la compréhension des relations de parenté entre les différents groupes de métazoaires a connu un renouveau ces 20 dernières années. Une des raisons est la mise au point de la méthode d'analyse cladistique, qui a pour principe de polariser les caractères observés par rapport à une référence externe au groupe d'étude, et à ne regrouper les organismes que sur la base de caractères dérivés partagés, les synapomorphies, et non pas sur la base d'une absence de caractères, comme c'était souvent le cas auparavant. Par exemple, s'il est tout à fait pertinent de regrouper ensemble tous les vertébrés parce qu'ils possèdent tous des vertebres, que l'on suppose apparues une seule fois chez leur ancêtre commun à tous, il n'y a pas plus de raison de grouper ensemble une moule, une fourmi et un oursin dans le groupe des « invertébrés » sous prétexte qu'il n'ont pas de vertèbres que de grouper Homme, fourmi et oursin à l'exclusion de la moule dans le groupe des « acoquillers » sous prétexte qu'ils n'ont pas de coquille. L'autre raison des progrès en matière de classification est due au développement de la microscopie électronique et de la disponibilité croissante de séquences génomiques, qui fournissent une grande quantité de nouveaux caractères utilisables pour des comparaisons.
Du fait de ces progrès, pour la première fois depuis 150 ans, un semblant de consensus se dégage au sujet des relations de parenté entre les grands groupes de métazoaires, et il peut être utile de connaître quelques grandes subdivisions. Celles-ci fournissent le cadre de base à partir duquel on peut discuter des questions relatives à l'histoire des métazoaires, par exemple: est-ce que les organismes proches constituent des « embranchements » anatomiquement homogènes, correspondant à un type de "plan d'organisation" particulier, et si non jusqu'où va la diversité au sein d'un même groupe?
Le résumé qui suit vise à fournir un aperçu de la diversité des métazoaires, non exhaustif mais suffisamment riche pour permettre de répondre à cette question. Seuls les groupes « majeurs », c'est à dire importants en termes de nombre d'espèces décrites seront présentés en détail, et reportés sur la figure 1. pour faciliter l'orientation. D'autres groupes numériquement moins importants, parfois limités à quelques espèces, seront toutefois évoqués, lorsque certains caractères observables chez eux fournissent des éléments utiles à la discussion.
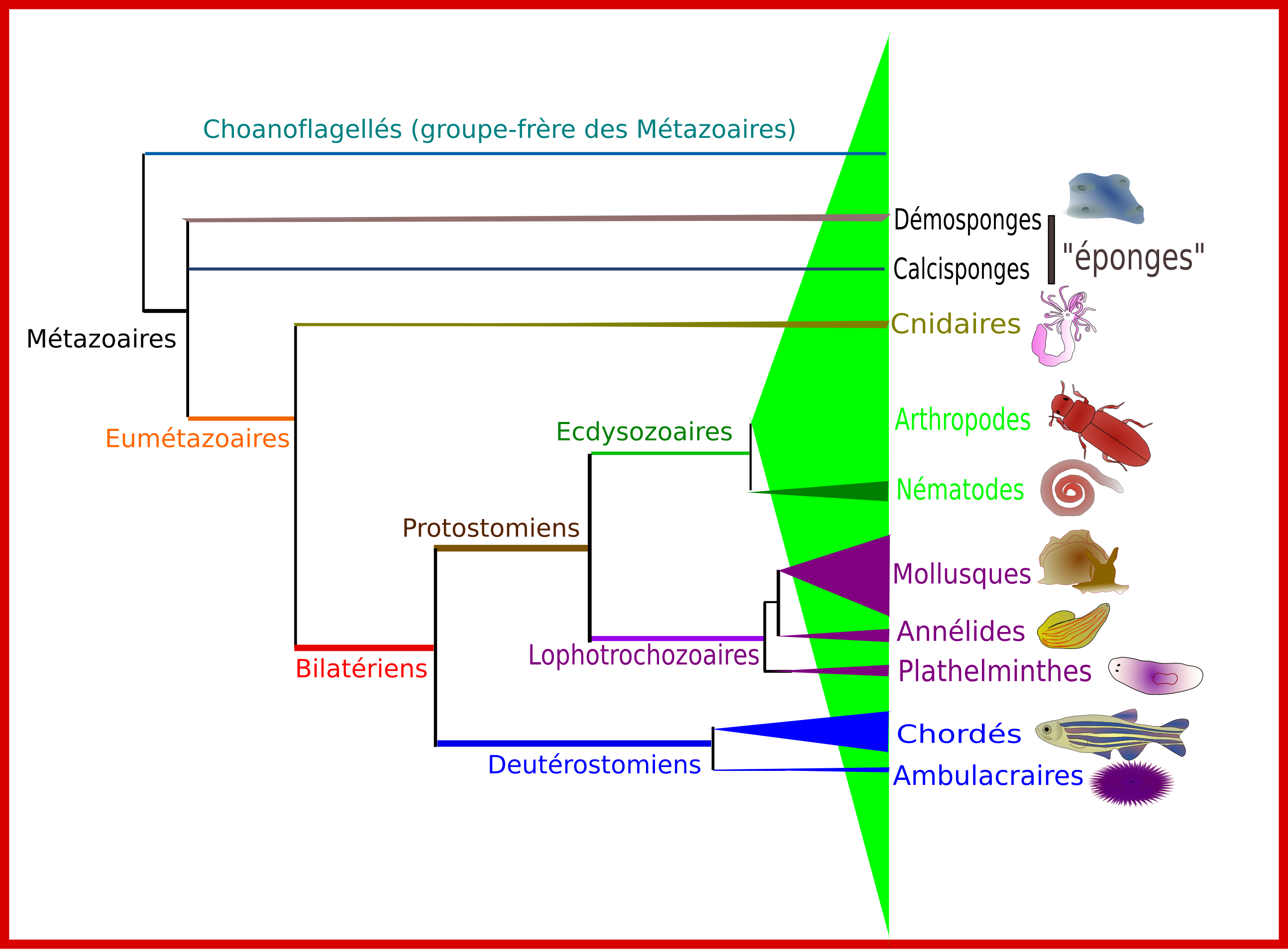
Figure 1: Relations de parenté entre les principaux groupes de Métazoaires.
Les longueurs de branches horizontales sont arbitraires. Les hauteurs des triangles sont proportionnelles au nombre d'espèces connues. Il convient toutefois de noter qu'il ne s'agit que du nombre d'espèces décrites, et que pour l'essentiel des groupes de petite taille et/ou difficiles d'accès, ces chiffres sont très sous-estimés.
 HAUT DE PAGE
HAUT DE PAGEII - La place des métazoaires parmi les être vivants
Les métazoaires ne constituent qu'un petit groupe parmi l'ensemble des êtres vivants (1 200 000 espèces décrites environ). S'il est légitime de s'y intéresser, étant donné ses facilités d'étude (organismes observables assez souvent à l'oeil nu) et son importance pratique (présence de nombreuses espèces d'intérêt agronomique ou médical, ainsi que de l'espèce humaine), il est important de prendre conscience qu'il ne s'agit là que d'une fraction infime de la biodiversité totale.
L'hypothèse de la monophylie des métazoaires ( c'est à dire du fait qu'ils descendent d'un ancêtre commun, propre uniquement aux membres de ce groupe) a été proposée par Haeckel dès 1877, et est soutenue par les analyses moléculaires. Le groupe frère des métaozaires est constitué par les choanoflagellés, un petit groupe d’eucaryotes unicellulaires (120 espèces connues). La cellule des choanoflagellés est caractérisée par la présence d'une colerette de microvillosités au centre de laquelle bat un flagelle, qui crée un courant d'eau acheminant les bactéries dont le choanogflagellé se nourrit au fond de la colerette (figure 2. A). Les choanoflagellés sont planctoniques ou fixés sur des algues. On les rencontre principalement en milieu marin, mais certaines espèces vivent en eaux douces. Les similitudes d'organisation entre les choanoflagellés (figure 2. A) et les choanocytes des éponges (fig 2.B et 2.C) sont interprétés comme une homologie, c'est à dire qu'il est supposé que ces structures étaient déjà présentes chez l'ancêtre commun des choanoflagellés et des métazoaires.
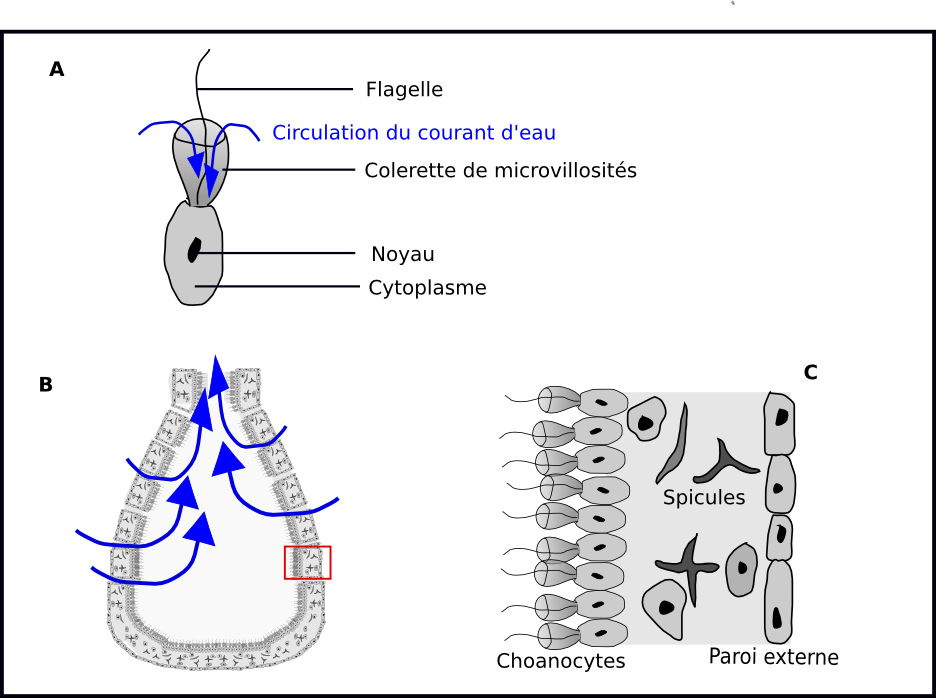
Figure 2: Choanocytes et choanoflagellés
A: Organisation d'un choanoflagellé libre. B: coupe transversale d'une éponge C: détail cellulaire de la paroi d'une éponge.
Par rapport aux choanoflagellés, parmi lesquels certaines espèces (du genre Proterospongia) sont coloniales, les métazoaires se distinguent par la présence de certains types de jonctions intercellulaires (jonctions septées et jonctions adhérentes), par la présence de molécules spécifiques de la matrice extracellulaires (collagène) et par un grand nombre de protéines régulatrices, notamment celles codées par des gènes à homéoboîte. Le génome des choanoflagellés contient des domaines protéiques impliqués dans la formation de la matrice et l'adhésion cellulaire chez les métazoaires, mais les combinaisons de ces domaines au sein des protéines sont différentes de celles rencontrées chez les métazoaires.
III - Les « éponges », un groupe en sursis...
Les « éponges » (10 000 espèces décrites) désignent un ensemble d'animaux caractérisés par la présence de choanocytes (voir figure 2.C), cellules flagellées à collerette, impliquées dans la nutrition, et d'un système aquifère, qui permet la circulation de l'eau à travers l'organisme. Du fait de leur mode de vie souvent marin (qui rend leur élevage difficile) et filtreur (qui provoque des risques importants de contamination par leurs proies en cas de séquençage génomique), ces animaux sont encore très mal connus d'un point de vue génomique, et leurs relations exactes avec les autres métazoaires sont encore sujettes à débat. Au dela de ces points communs, les « éponges » présentent une grande diversité d'organisation. Elles se divisent en deux grands groupes dont la monophylie est reconnue: les silicisponges (9000 espèces), qui possèdent un squelette constitué de spicules siliceux, et les calcisponges (1000 espèces), qui possèdent des spicules calcaires. La plupart des calcisponges présentent également une symétrie radiale. Les calcisponges sont exclusivement marines tandis qu'il existe des espèces dulçaquicoles parmi les silicisponges. Parmi les Silicisponges, certaines (les homoscléromorphes) possèdent un épithélium avec lame basale, ce qui est généralement considéré comme un critère définissant la présence de « tissus vrais ». Notons enfin qu'il existe des exceptions au mode de vie filtreur, telles que les éponges carnivores des genres Asbestopluma et Chondrocladia. Chez ces espèces carnivores, le système aquifère a régressé, et des spicules modifiés permettent la capture de petites proies.
 |
 |
| Une silicisponge: Acarnus erithacus | Une calcisponge: Ascandra pinus |
IV - Les eumétazoaires
Les eumétazoaires sont des animaux dépourvus de choanocytes, dont le corps est constitué de vrais feuillets, organisés en tissus, avec un épithélium pourvu d'une lame basale, c'est à dire un assemblage de glycoprotéines secrétées par les cellules éptihéliales et qui leur servent de point d'ancrage, et des cellules musculaires et nerveuses. Ils possèdent un tube digestif, et la régionalisation de leur corps le long de l'axe antéro-postérieur est régulée par des gènes Hox. Les deux principaux groupes d'eumétazoaires sont les bilatériens et les cnidaires.
IV-A Les bilatériens
Les bilatériens sont des animaux présentant deux axes de polarité, antéro-postérieur et dorso-ventral, qui définissent une symétrie bilatérale. Les embryons présentent un troisième feuillet, le mésoderme, qui se met en place lors de la gastrulation entre l'ectoderme et l'endoderme (voir figure 3). Ce feuillet est à l'origine des appareils circulatoire, excréteur, génital, et des muscles. La diversité des bilatériens actuels est telle qu'il est encore difficile de savoir si certains caractères, tels que la présence d'une cavité mésodermique (le coelome), d'une division du corps en unités anatomiques et fonctionnelles (les métamères) ou d'un système nerveux condensé étaient déjà présents chez l'ancêtre commun de tous les bilatériens ou si ces caractères sont apparus de façon convergente dans différents groupes. Seul un progrès dans les connaissances sur les groupes dits « mineurs », du fait du faible nombre d'espèces connues actuellement, permettra de trancher le débat. Si l'on excepte quelques groupes mal connus, dont la position phylogénétique fait encore débat, les bilatériens se divisent en deux groupes numériquement très inégaux, les deutérostomiens et les protostomiens.
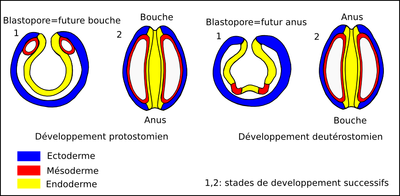
Figure 3: Développement embryonnaire des bilatériens.
Cette illustration très schématique de coupes longitudinales d'embryons à deux stades de développement successifs montre la différence entre un développement embryonnaire de type protostomien et un développement embryonnaire de type deutérostomien. Dans tous les cas, la mise en place des trois feuillets passe par l'invagination des cellules qui constitueront l'endoderme lors de la gastrulation. Comme l'endoderme est à l'origine du tube digestif, le blastopore, orifice par lequel s'effectue l'invagination, est à l'origine de l'une des extrémités de ce tube. Lorsque le blastopore est à l'origine de la bouche, le développement est dit « protostomien », et lorsque le blastopore est à l'origine de l'anus, le développement est dit « deutérostomien ».
La zone rouge représente le mésoderme, dont le creusement est à l'origine de la cavité coelomique.
IV - A - 1 Les deutérostomiens
Les deutérostomiens (58 000 espèces décrites) sont des animaux dont le blastopore est à l'origine de l'anus, mais ce caractère ne leur est pas propre, puisqu'il est également présent dans certains groupes de protostomiens (voir figure 3).
Les seules synapomorphies proposées actuellement pour le groupe seraient la présence de fentes pharyngiennes (voir figure 4), absentes chez les échinodermes, ce qui serait alors du à une perte secondaire. Les deutérostomes se divisent en deux grands groupes, les ambulacraires et les chordés, mais contiennent également l'énigmatique Xenoturbella, un petit animal benthique, pourvu d'un tube digestif à un seul orifice et d'un système nerveux diffus basiépithélial, qui se nourrit de larves de mollusques.
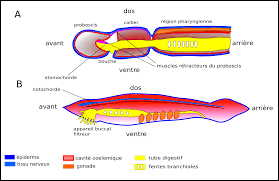
Figure 4: Comparaison de l'anatomie de deux deutérostomiens
A: Anatomie d'un entéropneuste (seule la partie antérieure de l'animal est représentée). B. Anatomie d'un céphalochordé.La présence de fentes pharyngiennes chez les deux animaux laisse penser qu'elles étaient présentes chez leur dernier ancêtre commun. Par contre, la "stomochorde" des entéropneustes et la notochorde des chordés sont des structures anatomiques très différentes: la stomochorde est un diverticule épaissi de la partie antérieure du tube digestif, tandis que la notochorde est une baguette rigide parcourant la partie dorsale du corps des chordés, en étroite association avec la chaîne nerveuse et les muscles locomoteurs. Malgré des noms qui prêtent à confusion, stomochorde et notochorde ne dérivent probablement pas d'une même structures ancestrâle, et sont apparues respectivement chez les entéropneustes et les chordés de façon indépendante.
IV-A-1-a Les ambulacraires
Les ambulacraires sont des organismes au corps divisé en trois régions dotées de trois systèmes coelomiques distincts. Au niveau moléculaire, il possèdent une particularité au niveau du code génétique mitochondrial, où le codon ATA code une isoleucine au lieu d'une méthionine. Ce clade contient les échinodermes (6000 espèces), très diversifiés morphologiquement et écologiquement (étoiles de mer, oursins, ophiures, holothuries, lys de mer), et particulièrement abondants dans les grands fonds, où ils constituent parfois l'essentiel de la biomasse. L'autre clade d'ambulacraires est celui des hémichordés (100 espèces), aux modes de vie et à l'anatomie également variés (entéropneustes psammivores et ptérobranches filtreurs). La présence d'un endostyle et de fentes branchiales chez les entéropneustes permet de supposer que ce caractère était présent chez l'ancêtre commun des deutérostomiens. Par contre, la "stomochorde", une structure rigide présente dans la partie antérieure de ces entéropneustes n'a rien à voir avec la notochorde (ou chorde), qui est une baguette cartilagineuse longitudinale permettant l'ancrage des muscles dorsaux chez les chordés (voir figure 4).
|
|
|
Un échinoderme : Protoreaster linckii |

IV-A-1-b Les chordés
Les chordés sont des animaux possédant une notochorde, une chaîne nerveuse dorsale et une hypophyse. Si l'on excepte les 11 espèces de céphalochordés, les 1200 espèces de tuniciers (ascidies, salpes, appendiculaires) et les 32 espèces de myxines, la grande majorité des membres de ce groupe sont des vertébrés, caractérisés par la présence de vertèbres, qui sont des pièces squelettiques qui entourent la chorde est se succèdent le long de l'axe antéro-postérieur. Les vertébrés constituent un groupe de 50 000 espèces, dont près de la moitié (23 000) sont des téléostéens (carpe, espadon, hippocampe, sole...), qui sont les vertébrés majoritaires dans les écosystèmes aquatiques, c'est à dire sur 98% du volume habitable du globe. Les tétrapodes, confinés pour la plupart dans les écosystèmes terrestres, se répartissent pour l'essentiel en 9600 oiseaux, 6850 squamates (lézards, varans, iguanes, serpents), 4900 lissamphibiens (grenouilles, tritons, cécilies) et 4500 mammifères.
IV-A-2 Les protostomiens
Ce groupe, défini sur des critères moléculaires, ne possède pas pour l'instant de synapomorphie morpho-anatomique. Notamment en ce qui concerne le développement embryonnaire, plusieurs groupes (chaetognathes, bryozoaires, phoronidiens) ont un développement de type deutérostomien, ceci permet de rappeler que le nom de protostome, comme tous les noms taxonomiques, a une fonction purement nominale et non descriptive. Les protostomiens constituent l'écrasante majorité des métazoaires (plus de 1 100 000 espèces), et se divisent, à quelques exceptions près, en ecdysozoaires et lophotrochozoaires.
IV-A-2-a Les ecdysozoaires
Les ecdysozoaires sont des animaux qui muent, mais il n'est pas démontré que la mécanique moléculaire qui déclenche ce phénomène est la même parmi les différents membres du groupe. Ils sont caractérisés par la perte des cils locomoteurs sur les cellules de l'épiderme, et la présence d'une cuticule en trois couches, dont la plus externe est secrétée par les microvillosités de l'épiderme. Les ecdysozoaires contiennent les arthropodes, les nématodes, et plusieurs autres groupes numériquement peu importants.
Ils constituent de loin le plus important groupe de métazoaires, avec près de 1 000 000 espèces décrites, dont 300 000 coléoptères, 150 000 diptères (mouches, moustiques), 125 000 hyménoptères (guêpes, abeilles, fourmis), 120 000 lépidoptères (papillons), 74 000 arachnides (araignées, acariens, scorpions), 12 000 myriapodes, autant de décapodes (crabes, crevettes) et de péracarides (cloportes, gammares). Présents dans tous les milieux, du fond des mers au sommet des arbres, ils sont caractérisés par la présence d'un squelette externe (ou exosquelette), subdivisé en pièces articulées entre elles. Tous possèdent également des appendices articulés (pattes, antennes, pinces....).
|
Un gastéropode: Chromodoris geometrica |

IV-A-2-b-3 Les plathelminthes
Les plathelminthes constituent un groupe de près de 14 000 espèces, caractérisées par un tube digestif présentant une seule ouverture (planaires) ou absent (ténia). Un grand nombre d'entre eux sont des parasites de l'homme ou du bétail (schistosomes, douves, ténias), mais d'autres sont des prédateurs libres, parfois terrestres. Certaines planaires ont des capacités de régénération particulièrement importantes.
 |
 |
 |
| Un cestode: Taenia saginatta | Une planaire: Pseudoceros dimidiatus | Un trématode: Schistosoma mansoni |
IV-B Les cnidaires
Ces animaux sont caractérisés par la présence de cnidocytes, cellules urticantes qui permettent la capture de leurs proies. Parmi les 10 000 espèces décrites, la majorité sont marines et prédatrices, mais il existe aussi des espèces symbiotiques, notamment parmi les coraux, et aussi quelques parasites, constituant le groupe des myxozoaires, qui s'attachent à leurs hôtes par des filaments interprétés comme des cnidocytes dérivés. Les cnidaires présentent une grande diversité des cycles de vie (phase polype ou méduse), des formes d'organisation (solitaire ou colonial, avec des architectures très variables). Parmi les anthozoaires (anémones et coraux), certaines espèces possèdent une symétrie bilatérale. Malgré un mode de vie fixé, de nombreuses espèces sont capables de se déplacer par mouvement de contraction de la colonne gastrique, et chez certaines hydres, il existe un orifice postérieur, maintenu ouvert activement et siège de transferts de matière. Les coraux batisseurs de récifs (scléractiniaires) ont une importance écologique majeure, puisqu'il sont à l'origine de nombreux biotopes marins.
|
|
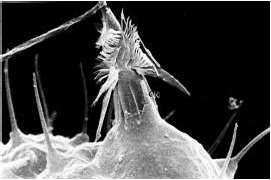 |
| Un scléractiniaire: Dendrogyra cylindrus | Un cnidocyte déchargé, observé au microscope électronique à balayage |
V - Conclusion
Quoique loin d'être exhaustif, cet aperçu de la diversité des métazoaires et de leurs relations de parenté permet quelques observations fondamentales:
- de nombreux caractères complexes (yeux camérulaires chez les vertébrés et les céphalopodes, segmentation du corps chez les vertébrés, pattes locomotrices chez les arthropodes et les vertébrés) sont présents uniquement chez des groupes très éloignés, et absent chez tous leurs apparentés. Ceci plaide en faveur d'une acquisition indépendante de ces caractères. L'absence de segmentation chez certaines annélides ou de tube digestif chez certains plathelminthes et chez Xenoturbella montrent que les pertes sont aussi des événements fréquents, et donc que l'évolution n'est pas dirigée vers une complexification permanente (ou plus exactement, que les pertes et les remaniements anatomiques qui en résultent sont une composante non-négligeable de cette complexification). Pour d'autres traits de caractères, tels que la condensation du système nerveux ou la présence d'un coelome chez l'ancêtre de tous les bilatériens, le débat reste ouvert.
- l'importance des taxons « mineurs » dans la discussion sur la robustesse de la classification met en évidence la nécessité de l'étude approfondie de la biodiversité pour comprendre l'histoire évolutive des métazoaires. Sans les entéropneustes, il ne serait pas possible de proposer de synapomorphies morpho-anatomiques pour l'ensemble des deutérostomiens, et certains caractères, probablement perdus chez les échinodermes, seraient alors interprétés à tort comme des innovations des chordés et non comme des marques de la condition primitive des deutérostomiens. Loin de constituer des « curiosités de la nature » anecdotiques, les membres des groupes dits mineurs fournissent (de même que les fossiles) des combinaisons de caractères différentes de celles présentes dans les « grands groupes », et permettent d'éviter les hypothèses simplistes (telle que celle de la complexification croissante) et les généralisations abusives qui reposent sur l'observation d'un nombre d'objets trop restreint.
- dans certains groupes, on observe une grande diversité anatomique, qui va à l'encontre des notions fréquemment véhiculées d' « organisme-type » ou de « plan d'organisation » supposé représentatif d'un groupe. Par exemple, les siboglidinés, les échiuriens et les siponcles étaient traditionnellement placés dans des embranchements distincts, alors qu'il sont désormais considérés comme des annélides très dérivés. Les myxozoaires ont subi le même sort en rejoignant les cnidaires. Il en fut de même beaucoup plus tôt avec les cirripèdes (balanes, sacculines), intégrés dans les arthropodes, alors que, inversement, le groupe des « vers » est depuis longtemps éclaté entre les plathelminthes, nématodes, annélides et autres organismes qui n'ont pas grand chose de plus en commun que l'absence de pattes locomotrices et de coquille.
Le désir de créer des entités d'importance particulière (tels que les embranchements) ou de déceler des tendances à la complexification dans une direction privilégiée relève de l'idéalisme, qui consiste à considérer la variation observable dans la nature comme un écart par rapport à un modèle idéal supposé représentatif. Si l'on considère à l'opposé que les organismes actuels sont le reflet d'une évolution permanente, qui suppose des pertes, des acquisitions ou des modifications de structures en réponse à la sélection exercée dans un milieu changeant, la diversité des métazoaires à tous les niveaux n'apparaît plus comme un casse-tête insoluble, mais comme une richesse, qui fournit des combinaisons d'états de caractères suffisamment variés pour permettre de retracer l'histoire des changements qui ont conduit à la diversification observable. L'étude de la classification phylogénétique apparaît alors non pas comme un exercice ésotérique et inutile, mais comme un cadre particulièrement adapté pour mettre en pratique les qualités d'observation et d'analyse méthodique de la réalité, qui constituent les principes du matérialisme scientifique, utiles même au delà des seules disciplines scientifiques.
Références
Lecointre, G. et Le Guyader, H. Classification phylogénétique du vivant, éd. Belin, 2006, 560 p.
Manuel, M., article « Evolution Animale », Encyclopedia Universalis (mis à jour en 2007).
Tree of Life, http://tolweb.org/ (projet de traitement exhaustif de la phylogénie des êtres vivants, avec références bibliographiques et discussions des incertitudes. Le degré de précision des informations est variable suivant les groupes.)