
Classification des végétaux
Introduction : de la difficulté de classer les végétaux
Classer les végétaux est un acte difficile, car la grande variété morphologique rend difficile de déterminer les caractères homologues, et en particulier l'état ancestral ou dérivé de ces caractères. De plus, certaines structures évoluent en fonction du développement du végétal. A l'heure actuelle, les comparaisons moléculaires tiennent une part importante dans l'établissement des phylogénies végétales. La classification des végétaux a ainsi grandement évolué pendant ces dernières décénnies.
Les difficultés de classement sont encore accentuées lorsqu'il s'agit de végétaux fossiles. Certains détails structuraux ne sont pas accessibles, les comparaisons moléculaires sont irréalisables.
De plus, les différentes parties d'un végétal (racines, tronc, feuilles, organes reproducteurs) sont généralement séparées donc fossilisées dans des endroits différents. Concrètement, ces différents fossiles ont été étudiés séparément, et se sont vu attribués des noms différents. En paléontologie, on parle ainsi de genre de forme déterminé pour un type d'organe précis (on parle aussi de genre d'organe). Plusieurs genres de forme peuvent ainsi correspondre en réalité à un même organisme. Cette situation de fait complique encore la détermination des relations de parenté entre ces espèces fossiles et les espèces actuelles. Par exemple, les arbres appartenant au genre Calamites se répartissent entre plusieurs genres de forme, dont : Calamites (au sens strict), pour les fossiles du tronc de l'arbre, et Annularia pour les rameaux.
|
Un tronc de Calamites : genre de forme Calamites |
des rameaux de Calamites : genre de forme Annularia |


La collection de Phylogène " forêt houillère " permet un classement utilisant les organes reproducteurs. Cette solution permet un classement relativement simple, même par des élèves de collège, et correspond à ce qui a longtemps été considéré comme étant l'évolution des végétaux terrestres. Les nouvelles classifications phylogénétiques présentent désormais des relations de parenté parfois différentes, basées en grande partie sur des comparaisons génétiques. Toutefois, les deux classifications ne sont pas incompatibles.
Arbre phylogénétique des Embryophytes
Les Embryophytes correspondent au groupe monophylétique regroupant l'ensemble des végétaux terrestres. Ils sont caractérisés par plusieurs caractères dérivés propres (synapomorphies) :
- présence d'un embryon pluricellulaire dans lequel plusieurs parties sont différenciées ;
- présence d'une cuticule ;
- gamètes formés dans des structures pluricellulaires, les gamétanges (gamétange femelle = archégone ; gamétange mâle = anthéridie) ;
- paroi des spores contenant de la sporopollénine, etc.
La classification des embryophytes a été grandement révisée depuis le milieu des années 1990. Par exemple, les anciennes Bryophytes polyphylétiques, ont été séparées en deux clades distincts, monophylétiques : les Marchantiophytes (hépatiques) et les Bryophytes (mousses s.s.).
Arbre phylogénétique simplifié des principaux clades d'Embryophytes
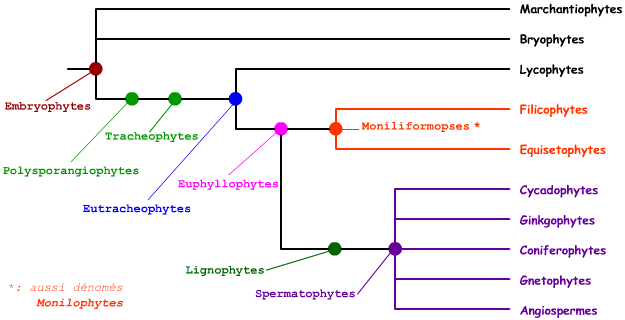
L'arbre présenté ci-dessus n'indique que les principaux clades d'Embryophytes : Il est donc grandement incomplet. Les points de couleur précisent les principaux groupes monophylétiques repérables ici. L'arbre étant volontairement simplifié, il ne faut pas s'étonner de voir parfois plusieurs noms successifs précisés pour une unique bifurcation de l'arbre.
A la base de l'arbre, on admet l'existence de plusieurs lignées distinctes, sans que leurs relations de parenté ne soient hiérarchisées dans l'état actuel des connaissances : les Marchantiophytes, les Bryophytes et les Polysporangiophytes (auxquels il faudrait rajouter les Anthocérorophytes, omis ici).
Les Marchantiophytes sont des végétaux de petite taille, terrestres, présentant une lame de tissu (thalle) ou un axe avec de petites feuilles. Il s'agit pour l'essentiel des hépatiques à feuilles. Les Bryophytes regroupent les mousses sensu stricto et les sphaignes. Il s'agit de petits végétaux, caractérisés entre autre par la présence d'un sporophyte chlorophyllien s'ouvrant par un opercule ou des valves, des feuilles non divisées...
Les Eutrachéophytes sont des Polysporangiophytes (plantes à sporophyte ramifié, pouvant porter plus d'un sporange), des Trachéophytes (présence de trachéides). En réalité, toutes les Polysporangiophytes actuelles sont des Eutrachéophytes. Au niveau de la flore actuelle, ces trois termes sont donc similaires, et il est légitime de n'employer que le terme d'Eutrachéophyte. Les espèces du genre Cooksonia (Silurien moyen, -425 millions d'années) sont les plus anciennes représentantes des Polysporangiophytes. Les Rhyniophytes (apparues il y a environ 420 millions d'années) sont les plus anciennes Trachéophytes, sans être toutefois des Eutrachéophytes.
Sur la base de critères liés au mode de croissance, les Eutrachéophytes sont divisées en deux clades : les Lycophytes (voir ci-dessous) et les Euphyllophytes. Ces derniers regroupent 99% des espèces actuelles de végétaux vasculaires. Une des caractéristiques principales des Euphyllophytes est la présence d'une croissance pseudomonopodiale : Alors que chez les autres Polysporangiophytes les deux rameaux présents à chaque bifurcation de l'axe ont une importance égale (croissance dichotome), un rameau présente une croissance verticale plus importante .
Les Euphyllophytes comportent deux clades distincts : les Moniliformopses (ou Monilophytes) et les Lignophytes. Cette distinction est établie essentiellement sur la base de données moléculaires, confortées par quelques données morpho-anatomiques. En particulier, le xylème (tissus conducteur de la sève brute) des Moniliformopses se développe à la fois vers l'intérieur et vers l'extérieur : on parle de xylème mésarche.
Les Moniliformopses comportent deux ensembles : les Equisétophytes (ou Sphénophytes, voir ci-dessous), qui forment un groupe monophylétique, et les Filicophytes (ou " fougères ", voir ci-dessous). Les Filicophytes forment en réalité un groupe paraphylétique.
Les Lignophytes sont caractérisées par la présence d'un cambium bifacial, produisant xylème et Phloème secondaire. Toutes les Lignophytes actuelles sont des Spermatophytes, caractérisées par la présence d'un ovule. L'ovule est une structure femelle comportant des tissus diploïdes (sporophytiques) et haploïdes (le -ou les- gamétophyte femelle) (pour les termes de sporophyte et de gamétophyte et leur place dans le cycle de développement des Spermatophytes, voir le document correspondant).
On distingue au moins cinq lignées distinctes au sein des Spermatophytes. La position relative de ces lignées est toujours grandement discutée à l'heure actuelle. En l'absence d'un consensus clair, il paraît raisonnable de ne pas chercher à hierarchiser les relations de parenté entre ces cinq lignées. Les Cycadophytes et les Gingkophytes (voir ci-dessous) possèdent un ovule, sans que l'ovule fécondé ne soit considéré comme une "vraie graine" : on parle parfois de pré-graine (remarque : cette distinction n'est pas faite par les Anglo-saxons, qui parlent de graine chez tous les Spermatophytes). Les Cycadophytes (apparues au Cambrien, il y a 270 à 280 millions d'années) ne sont plus représentées que par une centaine d'espèces, dont les Cycas. Les ovules sont portés par des feuilles très modifiées.
Les Spermatophytes à "vraie" graine comportent les Gnétophytes, les Coniférophytes (voir ci-dessous), et les Angiospermes (voir ci-dessous). Les Gnétophytes possèdent certains caractères de Coniférophytes et certains caractères d'Angiospermes. Elles ne comportent qu'environ 70 espèces, dont Ephedra distachya par exemple en France (dans les zones dunaires) et Welwitschia mirabilis, plante spectaculaire du désert du Namib, présentant deux uniques longues feuilles et pouvant vivre plusieurs centaines d'années.
Les Gymnospermes correspondent aux Cycadophytes, aux Ginkgophytes, aux Coniférophytes et aux Gnétophytes. Cet ensemble n'est pas monophylétique et il paraît donc maladroit de l'utiliser. Les Gnétophytes et les Angiospermes sont parfois regroupées dans l'ensemble des Anthophytes, sur la base de certains points communs importants (comme la double fécondation) ; ce regroupement est toutefois vivement contesté par certains auteurs, en particulier sur la base de données moléculaires. De même, Les Ginkgophytes et les Coniférophytes (qui sont alors dénommées Pinophytes) sont parfois regroupés dans un ensemble nommé Coniférophytes ; c'est en particulier le cas dans l'ouvrage de G. Lecointre et H. Le Guyader. La multiplicité de ces opinions encourage vivement les non-spécialistes que sont les enseignants du secondaire à ne pas choisir d'hypothèse dans l'immédiat, et donc à ne pas chercher à résoudre les relations de parenté précises entre les cinq clades de Spermatophytes.
Les groupes de la collection "forêt houillère"
Six clades d'embryophytes sont présents dans la collection "forêt houillère":
- les Lycophytes (Sélaginelle, Sigillaria)
- les Filicophytes (Polypode, Psaronius)
- les Equisetophytes (Prêle, Calamites)
- les Ginkgophytes (Ginkgo, Neuropteris)
- les Coniferophytes (Pin, Walchia)
- les Angiospermes (Chêne, Roseau)
Les Lycophytes regroupent tout un ensemble de végétaux actuels de petite taille. Les premiers représentants de cette lignée datent du Silurien supérieur. Elles présentent des sporanges solitaires, portés à l'aisselle de feuilles disposées autour de l'axe principal. Selon les espèces, les spores peuvent être toutes identiques (homosporie) ou bien être de deux tailles différentes (hétérosporie). L'exemple choisi pour la collection " forêt houillère ", la Sélaginelle, est hétérosporée. Si les Lycophytes actuelles sont de petite taille, ce n'était pas obligatoirement le cas par le passé : Dans la forêt houillère, nombre d'arbres de belle taille étaient ainsi des Lycophytes. Il s'agissait en particulier des Lepidodendron et des Sigillaria. (C'est ce dernier genre qui est présent dans la collection " forêt houillère ", mais le genre Lepidodendron aurait tout aussi bien convenu : Il est ainsi possible de faire effectuer des recherches complémentaires par les élèves sur cet arbre, de nombreux documents étant disponibles, tant sur Internet que dans la littérature).
Les Filicophytes correspondent aux Moniliformopses non-Equisétophytes. Il s'agit d'un ensemble paraphylétique. La majorité des espèces de Filicophytes (en particulier en France métropolitaine) correspondent en fait aux fougères leptosporangiées, qui forment elles un groupe monophylétique.
Les deux espèces de Filicophytes présents dans la collection " forêt houillère " sont :
- le Polypode, qui est une fougère leptosporangiée ;
- Psaronius, qui est une fougère arborescente, qui fait partie de la famille des Marattiacées. Il ne s'agit donc pas d'une fougère leptosporangiée.
Il est donc nécessaire d'utiliser le groupe des Filicophytes, bien qu'il ne soit pas monophylétique, pour regrouper ici le Polypode et Psaronius.
Les Filicophytes sont caractérisées entre autre par des feuilles, nommées frondes, qui se développent en crosses initialement.
Les Equisetophytes forment un groupe homogène regroupant les prêles actuelles et les Calamites du Carbonifère. Les rameaux sont insérés en verticilles sur la tige, ce qui correspond à une tige d'aspect articulé.
Les Ginkgophytes ne sont représentées actuellement que par une unique espèce, le Ginkgo biloba. Les plus anciennes Gingkophytes sont datées du Permien.
Dans la collection " forêt houillère ", le Ginkgo est rapproché de Neuropteris, bien que Neuropteris ne soit pas un Ginkgophyte. Ces deux plantes présentent des ovules sans qu'une graine se développe : Il ne s'agit donc ni de Coniférophytes ni d'Angiospermes.
Les Coniférophytes, ou Pinophytes, regroupent des Spermatophytes avec des cônes formés d'écailles. Ces écailles portent soit des ovules sur leur face supérieure, soit des sacs polliniques sur leur face inférieure.
Les Coniférophytes sont pour l'essentiel des arbres à feuilles persistantes en forme d'aiguilles ou de lanières. Plusieurs Coniférophytes étaient présents dans la forêt houillère : les Cordaïtales étaient les plus abondantes, mais Walchia a été choisi dans le cadre de la collection Phylogène, car plus proches des Coniférophytes Pinacées (auquelles appartient le Pin).
Les Angiospermes, ou plantes à fleurs et à fruits, sont caractérisées par une double fécondation avec siphonogamie, des vaisseaux vrais dans le xylème, des tubes criblés et cellules compagnes dans le phloème, etc. Elles sont apparues vraisemblablement peu avant le Crétacé (- 135 millions d'années) et ont rapidement évolué, dominant les flores terrestres dès la fin du Crétacé. Les deux espèces proposées dans la collection "forêt houillère" sont donc des plantes actuelles, les Angiospermes n'existant pas au Carbonifère ni au Permien :
- le Chêne est un arbre de la famille des Fagacées ;
- le Roseau est une plante herbacée monocotylédone de la famille des Poacées.
(Remarque : les monocotylédones, au contraire des dicotylédones, forment un ensemble monophylétique).
Quelques liens
Plusieurs sites Internet proposent de parcourir les classifications du monde vivant, et en particulier celle des Embryophytes. Voici quelques-uns de ces sites.
- [anglais] Page "Embryophytes" du serveur de Taxonomie du NCBI. Cette page permet de suivre l'ensemble de la systématique des Embryophytes. http://www.ncbi.nlm.nih.gov/Taxonomy/Browser/wwwtax.cgi?lin=s&p=has_linkout&id=3193
Il ne faut pas oublier les livres consacrés à la classification du vivant ou traitant en partie de la classification des végétaux :
- "Classification phylogénétique du vivant", 4ème édition (2016), Guillaume Lecointre, Hervé Le Guyader et Dominique Visset, Belin Ed. (584 pages)
- "Botanique : Biologie et Physiologie végétales" 3ème édition (2019), Sylvie Meyer, Catherine Reeb et Robin Bosdeveix, Maloine Ed. (608 pages)