
Différentes voies visuelles
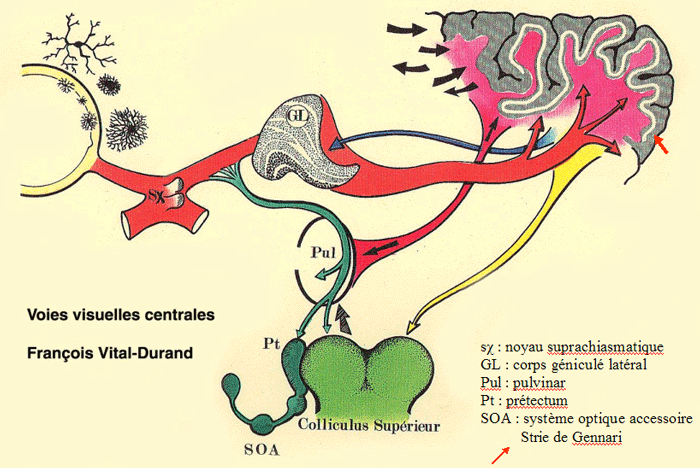 |
| Figure 1. Les voies visuelles centrales |
Les axones des cellules ganglionnaires de la rétine empruntent le nerf optique et se rendent directement au chiasma optique, à la base du diencéphale. Chez l'homme, environ 60% des fibres croisent dans le chiasma tandis que les 40% restants continuent du même côté vers le thalamus et le mésencéphale. Au dela du chiasma, les axones des cellules ganglionnaires forment le tractus optique (ou bandelette optique) qui, contrairement au nerf optique, contient des fibres provenant des deux yeux.
Ces axones pourront atteindre quatre cibles
-
dans le diencéphale: le corps genouillé latéral (CGL) du thalamus (diencéphale) puis, par les radiations optiques, le cortex visuel primaire: c'est la voie rétino-géniculo-striée ou "voie visuelle primaire", responsable de la majeure partie de la perception visuelle consciente
-
entre le thalamus et le mésencéphale: la région du prétectum (Pt), assurant la voie afférente du réflexe pupillaire à la lumière, "voie photique"
-
dans le diencéphale: le noyau supra-chiasmatique (sX) de l'hypothalamus, formant la "voie rétino-hypothalamique"
-
dans le mésencéphale: le colliculus, qui coordonne les mouvements de la tête et des yeux, et le pretectum, qui stabilise l'image, formant la "voie rétino-tectale"
La voie optique sensorielle primaire
En quittant l’œil pour le cerveau, on découvre un nouveau domaine d’application, des méthodes et des langages différents développés par les psychologues d’une part, et les neurobiologistes d’autre part. Pour la voie visuelle primaire, les psychologues parlent souvent de canaux spécialisés pour l’analyse de la forme, du mouvement et de la couleur, comme si chacune de ses modalités visuelles cheminaient le long de voies distinctes. Si ce paradigme des "canaux" permet d’organiser la pensée et la compréhension du phénomène de la vision, il décrit incomplètement le fonctionnement du système visuel : en effet, certains types de cellules ganglionnaires peuvent intervenir à la fois dans la reconnaissance des formes et le mouvement. Voir le traitement cérébral de l'information visuelle.
La voie rétino-géniculo-corticale
Notre perception consciente du monde visuel transite par des cellules ganglionnaires issues de la rétine et qui se projettent sur les corps (ou noyaux ) géniculés latéraux (CGL) droit et gauche.
Cette voie représente le devenir de 90% des fibres du nerf optique, soit 1,5 million d’axones des cellules ganglionnaires de la rétine se projetant sur le corps géniculé latéral (noyau visuel du thalamus) et du corps géniculé latéral au cortex visuel primaire (aire 17 ou V1).
Voir l'organisation des voies visuelles et la rétinotopie
De chaque côté du cerveau, le CGL reçoit l’information des cellules ganglionnaires codant l’hémichamp visuel hétérolatéral. L’hémichamp visuel gauche se projette sur le cortex strié droit par le biais d’informations provenant des deux yeux.
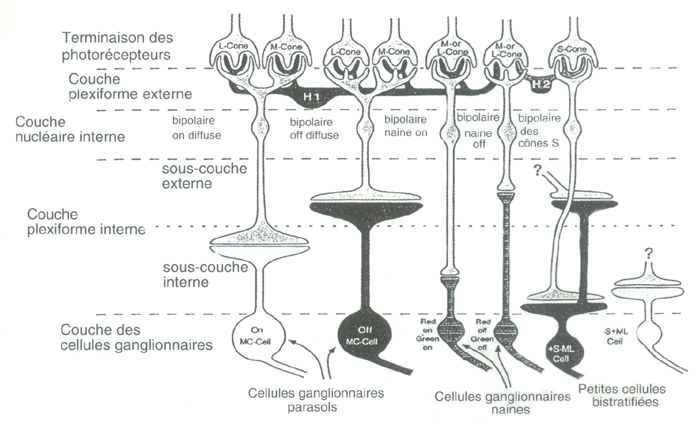
Le CGL est constitué de six couches superposées. Chaque couche reçoit l’information de l’hémichamp rétinien d’un seul œil. Les deux couches inférieures contiennent de gros corps cellulaires et constituent la zone magnocelluaire M (du latin magnus = gros). Les quatre couches supérieurs sont composées de cellules à corps plus petit et constitue la zone parvocellulaire P( du latin parvus =petit). Chaque couche présente en plus une sous-couche de corps cellulaires très petits appelée sous-couche koniocellulaire K (du grec konios = poussière). Les terminaisons axonales des cellules ganglionnaires qui se projettent sur chaque couche forment une carte rétinotopique précise : les cartes de toutes les couches sont superposées en correspondance exacte, de façon qu’un point donné du monde visuel perçu par les deux yeux se projette sur une ligne de projection traversant toutes les couches.
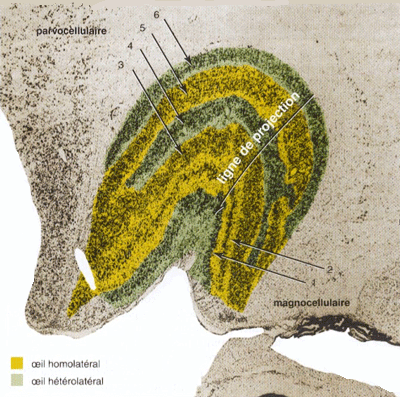 |
Figure 2. Organisation du corps (noyau) géniculé (genouillé) latéral Chaque CGL , droit ou gauche, reçoit des informations de l'hémichamp contralatéral, en provenance des deux yeux (rétine temporale ipsi-latérale et rétine nasale contra-latérale) ordonnées de façon rétinotopique. Par ailleurs, les différentes types de cellules ganglionnaires rétiniennes, constituant le début des 3 grandes voies M,P et K, y font relai dans des couchces séparées.
|
Les 3 voies fonctionnelles de la voie rétino-géniculo-striée
Il existe trois grandes voies fonctionnelles conduisant l'information visuelle de la rétine au cortex.
Les axones de toutes les cellules ganglionnaires naines (beta) se projettent uniquement sur le CGL, dans les portions principales des couches parvocellulaires (P). Ils font synapse avec une seule cellule P du CGL laquelle se projette sur la couche 4Cbeta du cortex strié dans la bande de dominance oculaire adéquat. Dans la fovéa, chaque cellule ganglionnaire naine reçoit l’information d’un seul cône au sein de la matrice des photorécepteurs.
Les voies de projection des cellules ganglionnaires parasol (a) présentent des caractéristiques assez proches de celles des cellules naines. Leurs axones se projettent, selon l’œil, dans l’une ou l’autre des sous-couches magnocellulaire (M). Leurs cellules relais du CGL se projettent sur la couche 4Calpha des bandes de dominance oculaire appropriées.
Les petites cellules ganglionnaires bistratifiées (gamma)se projettent sur les sous-couches K du CGL, et les cellules relais du CGL semblent se projeter au sein d’un lobule (ou colonne, ou blob) de la couche 3C
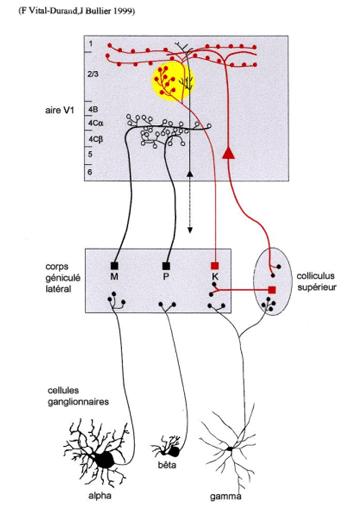 |
 |
|
Figure 3. Organisation des voies M, P et K issues de la rétine Les cellules de la rétine qui se connectent aux couches magnocellulaires du CGL sont les cellules ganglionnaire de type parasol (alpha), celles qui se connectent aux couches parvocellulaires sont les cellules ganglionnaires naines (beta), et celles qui se connectent aux couches koniocellulaires sont les cellules ganglionnaires bistratifiées (gamma) Au dela de l'aire visuelle primaire V1, la voie visuelle primaire se continue vers d'autres aires visuelles dites extrastriées qui traitent l'information à un niveau plus élevé. |
|
Spécialisation fonctionnelle des voies M, P et K
Bien que la correspondance ne soit pas stricte entre les petites cellules ganglionnaires rétiniennes (cellules béta, ou naines, ou Y) et les petites cellules du corps genouillé latéral (couche parvo-cellulaires), on oppose, dans un but simplificateur, les caractéristiques des voies Parvo-cellulaires (P) à celles des voies Magnocellulaires (M).
Les voies M sont sensibles aux contrastes de luminance, mais peu sensibles aux contrastes colorés. Elles saturent pour un contraste relativement faible (de l'ordre de 14%). Elle sont plus sensibles aux basses fréquences spatiales et aux fréquences temporelles élevées.
Les voies P sont peu sensibles aux contrastes de luminance et saturent à des niveaux élevés (elles présentent un gain faible pour le contraste). Elles sont très sensibles aux contrastes chromatiques, aux fréquences saptiales élevées et temporelles basses.
Il est possible avec des stimuli adaptés de séparer ces deux voies lors d'analyse électrophysiologiques par PEV: et pour
-
pour isoler l'activité des voies P, on utilise des stimulations de haute fréquence spatiale, de préférence avec des contrastes colorés, couvrant un petit champ visuel central, à basse fréquence temporelle
-
pour tester les voies M, on utilise des stimulations de basse fréquence spatiale, achromatique, de contraste faible (environ 15%) et couvrant un large champ visuel
|
Tableau récapitulatif des trois systèmes M,P,K |
RETINE |
CGL |
Physiologie |
Fonction |
|
P parvosystème |
Naines (X), bêta, B 80% |
couches 3 à 6 |
petit champ récepteur bonne acuité sensible hautes fréquences spatiales et basses fréquences temporelles couleur lent (tonique) |
assurerait la vision des détails et des couleurs |
|
M magnosystème |
Parasols (Y), alpha, A 10% |
couches 1 et 2 |
champ récepteur moyen acuité médiocre sensible basses fréquences saptiales et haute fréquences temporelles achromatique rapide (réponse phasique) |
assurerait la vision du mouvement et du papillotement
|
|
K koniosystème |
(W), gamma 10% |
interlaminaire |
grand champ récepteur couleur? réponses variées lent ou rapide |
? |
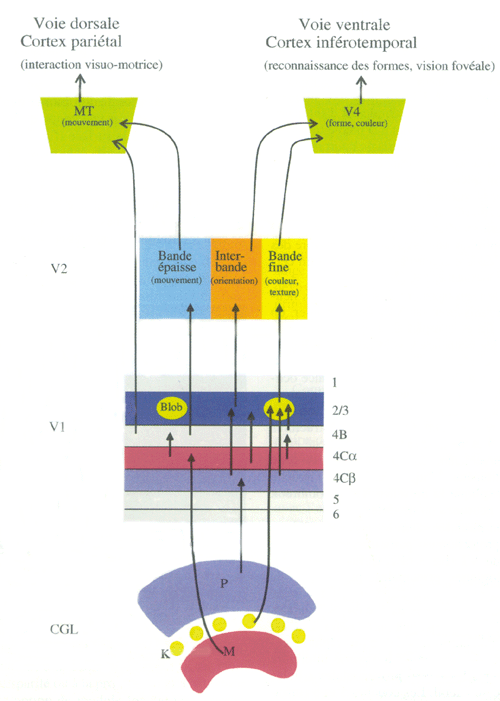 |
| Figure 4. Organisation fonctionnelle de la voie visuelle primaire (voir Les voies afférentes et le traitement de l'information) |
Les voies optiques secondaires et accessoires
La voie photique: la régulation des rythmes circadiens
Projection des cellules ganglionniares sur les noyaux suprachiasmatiques.
Ces noyaux sont présents chez tous les vertébrés. Ils sont situés au dessus du chiasma optique. Ces deux noyaux sont placés dans la ligne médiane du cerveau et reçoivent les axones des cellules ganglionnaires émergeant du chiasma optique. Ensemble ils synchronisent le rythme circadien de l’organisme. Celui-ci influe sur de nombreux aspects de notre physiologie : variations journalière de la température, sécrétions hormonales, veille et sommeil. Selon toute vraisemblance, les noyaux suprachiasmatiques reçoivent les projections axonales de cellules ganglionnaire recevant directement l’information des bâtonnets, et court-circuitant l’action des cellules bipolaires et amacrines (qui amplifient les différences d’intensité lumineuse aux dépens des niveaux absolus).
La voie rétino-tectale: l’usine oculomotrice
Projection de 10% des axones du nerf optique vers le colliculus supérieur (programmation des saccades oculaires), et le prétectum (stabilisation de l’image sur la rétine).
Le colliculus intègre les informations visuelles et auditives aux mouvements de la tête, et dirige les yeux vers les centres d’intérêt du monde extérieur. C’est la région visuelle la plus important du cerveau chez les poissons, les amphibiens et le reptiles : elle est appelée tectum. C’est encore une région majeure du cerveau des oiseaux. Chez le mammifères, le néocortex est devenu le lieu de la majorités des fonctions visuelles, mais le colliculus supérieur conserve la fonction essentielle de diriger les yeux vers les centres d’intérêt.
Le colliculus supérieur reçoit des informations de types très variés de cellules ganglionnaires. Leur nombre reste inconnu du fait des difficultés à les distinguer nettement sur des seuls critères morphologiques.
Le prétectum est situé au milieu des axones des cellules ganglionnaires qui se projettent sur le colliculus supérieur. A ce stade de la projection, ces axones sont groupés en fonction de leur destination rétinotopique au sein du colliculus supérieur. Certains de ces axones se terminent dans le prétectum (chez le chat, certaines cellules ganglionnaires se divisent pour se projeter à la fois sur le prétectum et le colliculus supérieur) Le prétectum joue un rôle dans l’ajustement de la taille de la pupille en réponse aux changements d’intensité lumineuse et dans le réflexe de poursuite des grands objets en mouvement.
Voir la partie relative aux mouvements oculaires.
Le système optique accessoire
Il doit son nom au fait que les faisceaux neuronaux qui le constituent quittent la voie de projection principale. Il est constitué de petits groupements cellulaires : noyau terminal dorsal, noyau terminal latéral, noyau terminal médian. Le système accessoire reçoit l’information d’un groupe de cellules ganglionnaires dites sélectives de direction de type ON : ces cellules répondent préférentiellement au mouvement dans unes direction spécifique. Il existe trois groupes de cellules répondant spécifiquement aux trois directions, et dont les axones rejoignent préférentiellement un des trois noyaux du système. Ces cellules ganglionnaires participent au maintien de la direction du regard pendant les mouvements de la tête, et donc à la stabilisation de l’image.
Voir la partie relative aux mouvements oculaires.